Старение и канцерогенез: равновесие на грани фола
МЕНЮ
Главная страница
Поиск
Регистрация на сайте
Помощь проекту
Архив новостей
ТЕМЫ
Новости ИИ
Городские сумасшедшие
ИИ в медицине
ИИ проекты
Искусственные нейросети
Искусственный интеллект
Слежка за людьми
Угроза ИИ
ИИ теория
Компьютерные науки
Машинное обуч. (Ошибки)
Машинное обучение
Машинный перевод
Нейронные сети начинающим
Психология ИИ
Реализация ИИ
Реализация нейросетей
Создание беспилотных авто
Трезво про ИИ
Философия ИИ
Генетические алгоритмы
Капсульные нейросети
Основы нейронных сетей
Промпты. Генеративные запросы
Распознавание лиц
Распознавание образов
Распознавание речи
Творчество ИИ
Техническое зрение
Чат-боты
Авторизация
2016-05-14 10:02
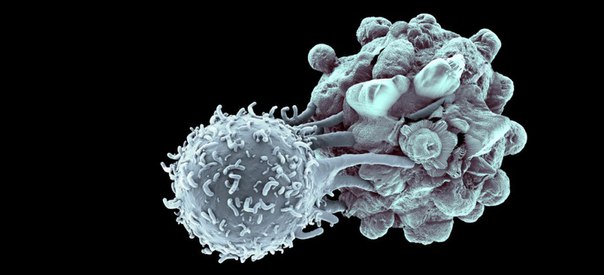

Биолог Константин Андреев рассказывает о жизненном цикле клетки, клеточной специализации и клеточной смерти; объясняет, в чём суть постулата Вейсмана, что такое лимит Хейфлика и почему клеточное бессмертие для организма - это очень плохо.
Вчера на улице ко мне подошла старуха и предложила купить вечную иглу для примуса.
Вы знаете, Адам, я не купил. Мне не нужна вечная игла, я не хочу жить вечно.
И. Ильф, Е. Петров «Золотой теленок»
Само понятие жизни и смерти как таковых у отдельной клетки и любых многоклеточных конструкций, будь то культура на донышке чашки Петри, участок эпителия, выстилающего легкое, 959-клеточная нематода Caenorhabditis elegans или в крайнем случае цельный человек, носит совершенно разный смысл. Для сложноорганизованных систем, к коим принадлежим и мы, определение «живой - неживой» формулируется довольно просто и базируется на полудюжине признаков, о которых студентам любых биологических факультетов рассказывается в течение их первого семестра обучения. И только наличие их всех в совокупности позволяет отнести изучаемый объект к живой природе.
С точки зрения физиологии ключевым из этих признаков должна быть способность живой особи к обмену веществом и энергией с окружающей средой, что включает в себя такие процессы, как питание, дыхание и выделение. С биохимической точки зрения живой системой, пожалуй, следовало бы назвать ту, внутри которой продолжаются реакции метаболизма и поддерживается постоянство ее химического состава. В понимании биофизиков жизнь - это извечная борьба с энтропией, точнее, умение поддерживать уровень беспорядка в системе ниже такового за ее пределами. Генетика определяет живой объект как тот, который обладает способностью к самовоспроизведению, благодаря чему все молекулы, клетки, ткани схожи по строению со своими предшественниками. С позиции эмбриологии жизнь подразумевается как конечный процесс онтогенеза, начиная с состояния одноклеточной зиготы и заканчивая моментом, когда все дальнейшие изменения и приобретение новых свойств завершаются. Причем качество этих изменений, будь то рост с накоплением биомассы или, напротив, отмирание тканей, роли не играет (то есть когда вам указывают на растущую лысину, можно это бойко парировать тем, что, мол, вы не стареете вовсе, а, можно сказать, развиваетесь). Этологи, в свою очередь, ставят во главу угла наличие у живого объекта раздражимости или избирательной реакции на внешние воздействия. Если ткнуть, скажем, обыкновенную амебу (Amoeba proteus) чем-нибудь острым, то она вполне благоразумно должна уползти прочь, а если, к примеру, добавить в среду что-либо питательное, то, напротив, немедленно начнет перемещаться по градиенту концентрации этого питательного в сторону его источника. Сходный эксперимент, кстати говоря, превосходно сработает и на спящем коллеге, разве что тыкать придется чем-то поувесистей, а роль аттрактивного фактора вместо питательной среды в данном случае лучше всего сыграет запах свежесваренного кофе. Наличие подобной избирательной реакции и дает основания предполагать, что перед нами живой организм.
Для отдельной клетки все выглядит несколько иначе. Начнем с того, что весь жизненный цикл для нее - это всего лишь временной промежуток между двумя митотическими делениями и что любой акт рождения новой клетки неразрывно связан со смертью ее предшественницы. С другой стороны, митоз, уже по природе своей предполагающий полную идентичность двух получившихся дочерних клеток, де-юре обеспечивает ей некое формальное бессмертие. В действительности, конечно, все происходит несколько не так, и в природе существует немало факторов, нарушающих правильное протекание митоза (попробуйте, к примеру, обработать клетку колхицином и посмотрите, что из этого выйдет). Да и в обычных условиях разновидностей клеточного деления, которые приводят к неравномерному перераспределению генетического материала между потомками, тоже хватает, включая кроссинговер при мейозе, амитоз, поли- и анеуплоидию, кариогенез, приводящий к формированию многоядерного синцития, и так далее.
Тем не менее любой из вышеперечисленных вариантов клеточного деления для ткани в целом будет служить основой для ее дальнейшего роста. Чтобы этот рост не вышел из-под контроля, существуют три вида настоящей клеточной гибели, широко описанные в научной литературе: аутофагия, некроз и апоптоз. Из всех них лишь последний является запрограммированным и точно управляемым процессом и потому ключевым игроком во всех тех процессах, о которых пойдет речь ниже.
Сразу хочу подчеркнуть, что любой организм существует в условиях непрекращающегося балансирования между делящимися и умирающими клетками - примерно так же, как бухгалтерия сводит дебит с кредитом, и для взрослого человека, чье тело в среднем состоит из 1013 клеток, ежесекундный их оборот составляет где-то около 106-107. Если это довольно хрупкое равновесие вдруг по какой-либо причине будет нарушено, то в зависимости от того, в какую из сторон оно будет смещено, на выходе мы будем иметь или опухолевый рост, или же омертвление тканей. Любопытно, но именно клеточная ткань либо клеточная культура как уровень организации живой материи в случае, когда интенсивность пролиферации клеток не компенсируется их гибелью, является единственным примером действительного бессмертия, что с успехом используется в современной науке. И эти единственные бессмертные клетки - рак.
Вечная жизнь Генриетты Лакс
Среди множества иммортализованных клеточных линий самая известная на сегодняшний день - это линия HeLa. Она была получена в 1951 году из эндотелиальных клеток шейки матки, взятых у раковой пациентки по имени Генриетта Лакс (HeLa - акроним имени и фамилии Henrietta Lacks) незадолго до ее смерти. Зараженные изначально вирусом папилломы, раковые клетки были подвержены хромосомным аберрациям и содержали вместо стандартных 46 человеческих хромосом 82. Однако другой, куда более любопытной и значимой их особенностью оказалась «вечность» - способность бесконечно делиться в лабораторных условиях. А когда обнаружилось, что клетки HeLa можно пересылать почтой и, более того, даже десятилетиями хранить в морозильной камере, а затем вновь продолжать культивировать на искусственной питательной среде, это тут же привело к громадному ажиотажу в научном мире, а линия HeLa немедленно разошлась по лабораториям во всех уголках земного шара. Впоследствии, впрочем, имел место серьезный скандал с их участием. Когда анализ хромосомных наборов ряда других клеточных линий, использовавшихся для исследований, выявил случаи их контаминации клетками HeLa, это привело к тому, что все результаты, полученные на основе экспериментов на этих линиях и уже опубликованные в научной печати, на деле не могли считаться достоверными. В итоге вышла немалая путаница, стоившая кому-то карьеры и репутации. Однако, несмотря на это, интерес к HeLa не охладел и до сих пор, поскольку раковые клетки с известной долей допущения являются довольно адекватной моделью для поиска ответов на многие биомедицинские вопросы. И именно клетки линии HeLa дали возможность Джонасу Солку создать вакцину против полиомиелита, поучаствовали в проекте клонирования знаменитой овцы Долли (предварительные опыты по пересадке ядер проводились как раз на HeLa), а также использовались для составления генетических карт, отработки искусственного оплодотворения и прочих поставленных научных задач. И даже слетали в космос в 1960 году в рамках космической программы советских генетиков.
Постулат Вейсмана и лимит Хейфлика
Что же именно делает раковые клетки бессмертными? И почему все остальные клетки подвержены тому, что мы зовем старением? И, в конце концов, что такое само старение? Первым, кто подошел к этому вопросу вплотную, был известный зоолог и теоретик эволюционного учения Август Вейсман, предложивший еще в 1881 году свой ставший впоследствии знаменитым постулат, который гласил, что у соматических клеток «-способность к росту путем деления не вечна, а ограничена», чем и обусловливается старение всего организма. Бесконечно же подвергаться делению, по его мнению, могли только половые клетки. Разумеется, все эти выводы были сугубо эмпирическими, так как ни о каких экспериментальных методах работы с живыми клетками в то время и речи быть не могло. Какое там! Ведь даже сама клеточная теория Шлейдена - Шванна о том, что вообще есть клетка, была сформулирована лишь тридцатью годами ранее.
Позднее постулат Вейсмана подвергался испытанию на прочность опытами французского хирурга и патофизиолога А. Карреля, который впервые применил на практике методику выращивания культуры тканей, выделенных из организма. Техника эксперимента была проста и вроде бы полностью опровергала вейсмановское предположение о так называемой «смертности» соматических клеток. Отрезанный кусочек миокарда куриного сердца, помещенный в питательную среду и инкубированный в термостате, разделялся затем на две равные части, которые пересаживались в новые, отдельные пробирки и инкубировались дальше. Подобные пересевы можно было продолжать в течение долгого времени, чуть ли не лет, и на протяжении всех этих пассажей фибробласты миокарда продолжали и продолжали делиться. За эти исследования Каррель в 1912 году был удостоен Нобелевской премии, хотя, как оказалось впоследствии, его эксперимент был выполнен не совсем-таки чисто. Виной тому была эмбриональная жидкость, использовавшаяся в качестве питательной среды для поддержания жизнедеятельности клеточной культуры: содержащиеся в ней свежие клетки, попадая в образец и активно делясь, создавали тем самым мнимый эффект бессмертия «старых» фибробластов, ибо что там разглядишь в пробирке!
И эксперимент был переделан полвека спустя американским цитологом Л. Хейфликом. В отличие от Карреля, он помещал в питательную среду не сам по себе кусочек извлеченной из организма ткани, а предварительно обработанный трипсином, благодаря чему ткань распадалась на отдельные клетки. Кроме того, и сам выбор питательной среды был другим: вместо эмбриональной сыворотки или плазмы крови Хейфлик использовал искусственно подобранный раствор из аминокислот, солей и прочих низкомолекулярных компонентов, достаточных, чтобы поддерживать клеточную репродукцию. В остальном метод был тот же: попадая в питательную среду, фибробласты немедленно начинали делиться, и, как только их слой достигал определенного размера, образец делился пополам, еще раз обрабатывался трипсином и пересевался в новую пробирку/чашку Петри. И после определенного числа пересевов, примерно соответствующего 50 клеточным делениям, рост ткани останавливался, а переставшие делиться клетки через некоторое время погибали. Это явление наступало регулярно, неотвратимо и наблюдалось во всех повторных опытах, впоследствии проведенных независимыми группами исследователей, причем не только для фибробластов, но и для любых иных типов соматических клеток. Критическое число делений, отведенных соматическим клеткам на их жизнь, получило название лимита Хейфлика, а эксперимент полностью реабилитировал теорию Вейсмана относительно смертности клеточных линий в частности и любых организмов в целом.
Теломеразная теория старения. Как обойти закон
Вернемся к особенностям раковых клеток. Им единственным удается преодолеть этот барьер. И происходит это за счет двух взаимосвязанных между собой факторов: с одной стороны, активации у них фермента - теломеразы, с другой стороны, блокировки механизмов апоптоза - программируемой клеточной гибели. Как примерно работает вся эта схема? Теломеры - это некодирующие участки хромосом, локализованные на их концах (то есть по две на каждую линейную хромосому) и состоящие из многократно повторяющихся (примерно 10-15 тысяч) коротких нуклеотидных фрагментов, к примеру, у большинства позвоночных это TTAGGG. Хотя известно, что никакой генетической информации теломерная ДНК не несет, тем не менее ее роль весьма существенна. Во-первых, при незапланированном воздействии экзонуклеаз (ферментов, «отщепляющих» концевые мононуклеотиды от полинуклеотидной цепи) теломеры принимают удар на себя, смысловая ДНК при этом не страдает, и, таким образом, негативный эффект минимизируется. Однако даже это не главное. Основная функция теломер - защищать кодирующую ДНК от потерь при укорочении хромосом в ходе их репликации. Проблема возникает из-за того, что любая ДНК-полимераза осуществляет синтез дочерней цепи только в строго определенном направлении - от 5
Телеграм: t.me/ainewsline
Источник: vk.com