Древний ствол: где в мозге зарождается сознание
МЕНЮ
Главная страница
Поиск
Регистрация на сайте
Помощь проекту
Архив новостей
ТЕМЫ
Новости ИИ
Городские сумасшедшие
ИИ в медицине
ИИ проекты
Искусственные нейросети
Искусственный интеллект
Слежка за людьми
Угроза ИИ
Компьютерные науки
Машинное обуч. (Ошибки)
Машинное обучение
Машинный перевод
Нейронные сети начинающим
Психология ИИ
Реализация ИИ
Реализация нейросетей
Создание беспилотных авто
Трезво про ИИ
Философия ИИ
Генетические алгоритмы
Капсульные нейросети
Основы нейронных сетей
Распознавание лиц
Распознавание образов
Распознавание речи
Творчество ИИ
Техническое зрение
Чат-боты
Авторизация
2024-12-01 11:46
В издательстве «Альпина нон-фикшн» вышла книга «Скрытый источник сознания: В поисках природы субъективного опыта» нейропсихолога Марка Солмса. Опираясь на элементарную физику живого, новейшие достижения в аффективной нейробиологии и психоанализ субъективного опыта, автор выдвигает принципиально новое решение трудной проблемы сознания. Публикуем фрагмент из главы, посвященной значению ствола мозга, имеющего чрезвычайно древнее эволюционное происхождение.
В своей новой книге Марк Солмс представляет революционную теорию, согласно которой сознание обусловлено чувствами. Его порождает не только кора головного мозга, отвечающая за интеллект и высшие психические функции, но также более древняя часть мозга, где зарождаются эмоции. Кора становится носителем сознания лишь настолько, насколько ее возбуждает ствол мозга. Это и есть тот самый «скрытый источник». Автор связывает воедино элементарную физику живого, новейшие достижения вычислительной и аффективной нейробиологии, статистическую физику, термодинамику и принцип свободной энергии К. Фристона, а также тонкости субъективного опыта, которыми традиционно занимался психоанализ. Пристальное внимание он уделяет субъективному опыту сотен пациентов с неврологическими расстройствами, наблюдение за которыми позволило приблизиться к более полному пониманию работы мозга.
Основное положение нейропсихологии, основанное на клинико-анатомическом методе, гласит, что если определенная психическая функция выполняется определенной областью мозга, то полное разрушение этой области должно приводить к полной потере этой функции. Как я уже продемонстрировал, когда речь заходит о сознании, кора проваливает этот тест. Но с кортикальной теорией сознания дело обстоит еще хуже: сознание полностью утрачивается при повреждении других областей мозга, причем эти повреждения могут быть совсем незначительными.
Более 70 лет назад физиологи Джузеппе Моруцци и Хорас Мэгун установили, что кошки теряют сознание после микроскопического надреза, отделяющего кору от ретикулярной (т. е. сетчатой) формации в стволе мозга. Этой структуре, по-видимому, около 525 млн лет, так как она имеется у всех позвоночных, от рыб до человека. Со времени открытия Моруцци и Мэгуна исследователи подтвердили, что у представителей самых разнообразных видов сравнительно мелкие повреждения этой структуры — в научной терминологии известной как ретикулярная активирующая система — вызывают кому. В частности, Дэвид Фишер и его коллеги недавно нашли у людей, перенесших инсульт ствола головного мозга, повреждение в крошечном (два кубических миллиметра) участке в верхней части покрышки варолиева моста, характерное именно для комы (рис. 1).

Существует два возможных объяснения этому. Первое — что упомянутая плотноупакованная структура ствола мозга и есть место зарождения сознания, сокровенный источник психики, средоточие ее сущности. Я придерживаюсь именно такого мнения, как и ныне покойный Яак Панксепп. Согласно второму объяснению — это что-то вроде электрического шнура для телевизора: вещь необходимая, но недостаточная и вряд ли способная дать необходимую информацию тем, кто хочет разобраться, как работает телевизор. Это общепринятая точка зрения.
Предположим, вторая версия верна. Тогда при стимуляции ствола мозга можно ожидать, что сознание будет включаться и отключаться. В крайнем случае его можно будет определенным образом ослабить, подобно тому, как экран телевизора гаснет при падении напряжения в сети. Не стоит ожидать, что кабель на ходу перепишет содержание текущей передачи. Тем не менее электрод, имплантированный в одно из ядер ретикулярной системы мозга 65?летней женщины (в целях лечения болезни Паркинсона), неизменно вызывал следующий удивительный результат:
Через пять секунд на лице пациентки отобразилась глубокая печаль. <…> Будучи по-прежнему в сознании, пациентка наклонилась вправо, стала плакать и вербально выражать чувства грусти, вины, собственной никчемности и безнадежности, например: «Я упала в собственных глазах, я больше не хочу жить, не хочу ничего видеть, слышать, чувствовать…» Когда ее спросили, почему она плачет, как будто испытывает боль, она ответила: «Нет, мне надоело жить, с меня хватит… Я больше не хочу жить, жизнь мне противна… Все без толку, всегда чувствуешь себя ничтожеством, мне страшно в этом мире». На вопрос, что ее огорчает, она ответила: «Я устала. Хочу забиться в угол… Конечно, я по себе плачу… Я безнадежна, зачем вам со мной возиться?» <…> Депрессивное состояние исчезло менее чем через 90 секунд после прекращения стимуляции. Следующие пять минут пациентка находилась в состоянии легкой гипомании, смеялась и шутила с исследователем, игриво дергая его за галстук. Весь это эпизод она помнила. Стимуляция [другого участка мозга, в который и планировалось вводить электрод] не вызвала подобной психической реакции.
У этой пациентки в анамнезе не было никаких психиатрических расстройств.
То же происходит при химической стимуляции или блокаде этих ключевых ядер ствола мозга. Большинство антидепрессантов — серотониновые стимуляторы — воздействуют на нейроны, клеточные тела которых расположены в области ретикулярной активирующей системы, именуемой ядрами шва (рис. 1).
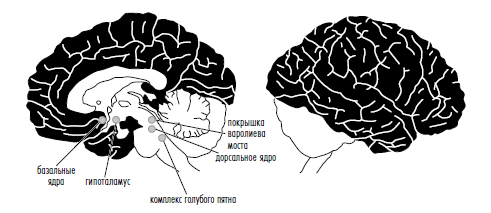
Это, как мы говорим, «поставщики» серотонина. Антипсихотики — блокаторы рецепторов дофамина — воздействуют на нейроны, расположенные в другой части ретикулярной активирующей системы: вентральной области покрышки (рис. 2). То же относится к успокоительным препаратам — многие из них блокируют норадреналин, химическое соединение, которое вырабатывается нейронами, расположенными в комплексе голубого пятна (рис. 1), еще одного элемента ретикулярной активирующей системы. Все эти нейроны сконцентрированы в ретикулярной системе ствола мозга. Психиатры не стали бы пытаться воздействовать на эту область мозга, если бы она просто включала и выключала сознание. Если бы ее роль ограничивалась только этим, она бы интересовала лишь анестезиологов. А следовательно, вторая точка зрения неверна.

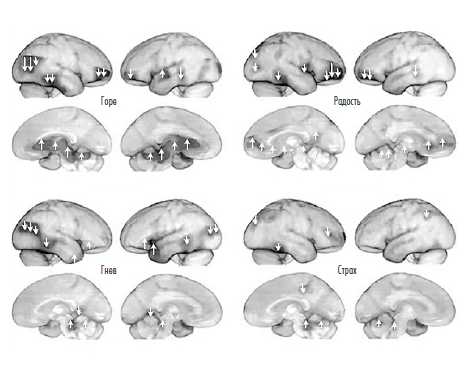
К тому же выводу приводят и результаты функциональной нейровизуализации мозга в различных эмоциональных состояниях. В частности, ПЭТ мозга людей в состояниях ГОРЯ, ПОИСКА, ГНЕВА и СТРАХА показывает наибольший уровень метаболической активности в ретикулярной системе ствола мозга (и других подкорковых областей, см. рис. 9), тогда как кора в это время деактивирована. То же показывают результаты фМРТ во время оргазма: повышение гемодинамической активности, коррелирующее с этим крайне аффективным состоянием, проявляется почти исключительно в среднем мозге.
Изучение повреждений, глубокая стимуляция мозга, фармакологическое воздействие и функциональная нейровизуализация — все это позволяет сделать такой же вывод: аффекты порождает ретикулярная система ствола мозга.
Следовательно, очевидно, что единственная известная нам часть мозга, необходимая для возникновения сознания в целом, оказывает столь же мощное воздействие на другую психическую функцию, то есть чувства. В предыдущей главе я показал, как чувства пронизывают весь сознательный опыт: каковы бы ни были прочие функции сознания, одна из центральных его задач, по-видимому, состоит в том, чтобы переживать чувства и реагировать на них (а чувства приходят изнутри и регулируют наши биологические потребности). Однако теперь уже можно предположить, что неврологические истоки аффектов и сознания как минимум на глубинном уровне тесно переплетены друг с другом, а может быть, на самом деле являются частями одного и того же механизма. Вопреки классическому представлению эмпиризма, согласно которому мы обретаем сознание через чувства, а также процитированному мною высказыванию Мейнерта, основанному на этом представлении, мозг, по-видимому, «излучает собственный жар».
Как же назвать этот базовый носитель, это таинственное вещество разума, как будто наполняющее нас изнутри? Нельзя назвать его состоянием бодрствования, по Земану, так как в этом случае придется признать сновидения разновидностью бодрствования, что абсурдно. Нельзя сказать, что сознание связано с количественным уровнем активности мозга, как считали Моруцци и Мэгун, так как вышеперечисленные факты демонстрируют, что оно обладает ярко выраженными качественными характеристиками.
Поэтому попробуем прибегнуть к третьему термину, используемому в литературе, — «возбуждение».
Это слово кажется мне подходящим, вполне нейтральным. И бодрствование, и сновидение связаны с возбуждением, и это понятие не исключает качественного аспекта, в отличие от слова «уровень». Более того, возбуждение определенно указывает на наличие чувств.
Но что такое возбуждение? Ранее мы обсуждали его только в свете поведения — например, мы рассматривали различия между комой, вегетативным состоянием и полностью ясным сознанием. Измеряется оно, как правило, по шкале комы Глазго, включающей проверку реакции открывания глаз у пациента, его вербальные ответы на вопросы и моторные реакции на указания и на боль. Однако возбуждение можно определять и физиологически.
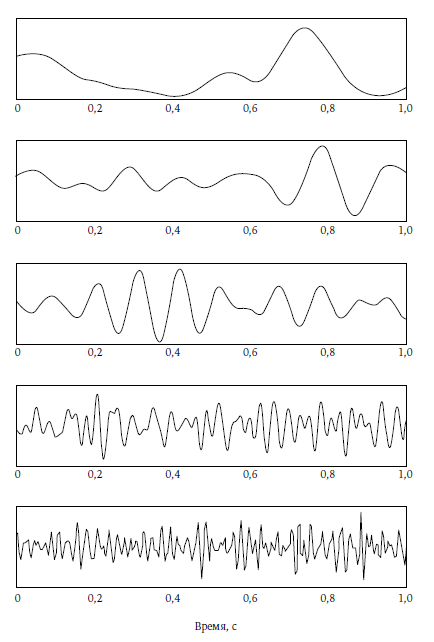
ЭЭГ дает графическое отображение электрической активности в коре мозга. Предоставленная самой себе (т. е. отключенная от ретикулярной активирующей системы даже во время обработки информации, поступающей от органов чувств), кора порождает дельта-ритмы — череду высокоамплитудных волн, возникающих примерно два раза в секунду (т. е. с частотой 2 Гц). При стимуляции ретикулярной активирующей системы в отсутствие сенсорной информации кора обычно порождает тета-ритмы (4–5 Гц) 5 или альфа-ритмы (десинхронизированные, т. е. нерегулярные волны с частотой 8–13 Гц). При активной обработке внешней информации в коре обычно наблюдаются бета-ритмы (десинхронизированные низкоамплитудные волны с частотой 14–24 Гц) или гамма-ритмы (низкоамплитудные волны с очень высокой частотой 25–100 Гц). Гамма-ритмы чаще всего принято связывать с сознанием.
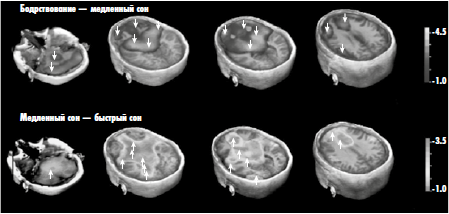
В наши дни физиологическое возбуждение можно измерить также с помощью методов функциональной нейровизуализации, которые позволяют в буквальном смысле создавать изображения активности мозга, отражающие паттерны изменений метаболизма в разных его участках. Эта технология была проиллюстрирована на рисунке 3 применительно к разным фазам сна. В нижнем ряду рисунка показано возбуждение во время быстрого сна, обычно (хотя, как было сказано выше, не обязательно) связанное с состоянием сознания во время сновидений и возникающее в верхней части ствола мозга. Нейровизуализация некоторых базовых эмоциональных состояний сознания (рис. 9) и оргазма демонстрирует то же самое — возбуждение в стволе мозга.
Кора становится носителем сознания лишь настолько, насколько ее возбуждает ствол мозга. Отношения между этими двумя структурами иерархичны: кортикальное сознание зависит от возбуждения ствола.
Вот почему дельта-ритм (рис. 10, вверху), не связанный с осознанным поведением, порождается сложной активностью коры, и вот почему гамма-ритм (рис. 10, внизу) можно стимулировать с помощью одной только ретикулярной активирующей системы. По этой же причине падение уровня физиологического возбуждения в верхнем ряду на рисунке 3 совпадает с угасанием сознания и засыпанием, а рост возбуждения в нижнем ряду совпадает с возвращением сознания при сновидениях. Эти факты бесспорны.
Давайте более глубоко рассмотрим задействованные при этом реальные мозговые механизмы. Для этого мне понадобится объяснить основное различие между двумя способами взаимодействия нейронов. Это различие, как выясняется, важно для сознания.
Синаптическая передача целенаправленна, бинарна («да/ нет») и протекает быстро. Этот аспект работы мозга больше всего напоминает цифровые вычисления, и, может быть, поэтому им так увлекаются представители вычислительной нейронауки. Этот процесс происходит по всей нервной системе, включая кору. Однако кора не является носителем сознания по определению. Иными словами, этот тип нейронной передачи осуществляется в коре независимо от того, является ли она источником сознания или нет. И она практически не имеет отношения к возбуждению.
Не столь широко известен тот факт, что синаптическая передача осуществляется под постоянным воздействием совсем иного физиологического процесса. Этот другой тип нейронной активности называется постсинаптической модуляцией.
В отличие от синаптической передачи, это процесс запутанный, явно химический и совсем не похож на то, что происходит в обычном компьютере. Он запускается эндогенно из ретикулярной активирующей системы (а также других подкорковых структур, и даже из некоторых органов, не входящих в нервную систему), как раз этот процесс имеет самое прямое отношение к возбуждению.
Главные участники этого процесса — молекулы, принадлежащие к классу так называемых нейромодуляторов. В отличие от нейромедиаторов, нейромодуляторы распространяются в мозге диффузно — то есть выделяются в относительной близости от целых популяций нейронов, а не на отдельных синапсах. Они не передают сообщения по конкретным каналам, а охватывают определенные участки сети, регулируя таким образом общее состояние коры. Например, состояния коры различаются на изображениях верхнего и нижнего рядов рисунка 3 (медленный сон / быстрый сон) и четырех эмоциональных состояний на рисунке 9 (ГОРЕ/ПОИСК/ ГНЕВ/СТРАХ). В каждом из этих состояний кора по-разному обрабатывает информацию. Так, например, если вас позовут по имени, когда вы спите, вы отреагируете совсем не так, чем тогда, когда вы бодрствуете. Аналогичным образом сравните вашу реакцию на приближение незнакомца, когда вы находитесь в состоянии ПОИСКА и когда — в состоянии СТРАХА: в первом случае вы можете обрадоваться незнакомцу и даже завязать с ним разговор, тогда как в последнем будете сторониться его в надежде, что он вас не заметит.
Проведение различия между передачей информации по каналам и регуляцией общего состояния коры полезно для понимания двух способов взаимодействия нейронов между собой. Состояние коры влияет на дифференциальные мощности передачи сигналов, проходящих по ее каналам; иначе говоря, оно регулирует «громкость» коммуникации по разным каналам (рис. 11). Вот почему один и тот же звук (например, когда кто-то зовет вас по имени) широко распространяется в коре при бодрствовании, но изолируется в пределах слуховой коры во сне, а при виде незнакомца запускается одна нейронная цепь в состоянии СТРАХА и другая — в состоянии ПОИСКА.
В этом и состоит суть того, что мы вкладываем в понятие «возбуждение». Отметим, однако, что возбуждение коры можно как усиливать, так и ослаблять вплоть до полного подавления передачи, что и происходит ночью, всякий раз, когда мы ложимся спать (поэтому некоторые физиологи предпочитают термин «модуляция», а не «возбуждение»). Возбуждение, следовательно, определяет, какие синаптические импульсы передаются и насколько мощно, как в приведенном примере с окликанием человека по имени во сне и в состоянии бодрствования.
Синаптическая передача бинарна («вкл/выкл», «да/нет», «1/0»), но постсинаптическая нейромодуляция регулирует вероятность того, что определенная комбинация нейронов даст импульс. Такая нейромодуляция меняет статистические шансы того, что с нейронами что-то произойдет. Эта вероятностная, аналоговая настройка частоты импульсов осуществляется через рецепторы, расположенные в нескольких местах вдоль всего нейрона. В отличие от нейромедиаторов, нейромодуляторы оказывают сравнительно медленное и долгосрочное воздействие — не только потому, что сами химические соединения существуют дольше, но и потому, что чем чаще импульсы идут по данному каналу, тем больше вероятность того, что этот канал будет и дальше их проводить. Если стимулировать участок нейронной сети, он останется несколько возбужденным до тех пор, пока модуляция не снизит возбуждение. Это влияет на нейропластичность и в немалой степени составляет основу обучаемости. Благодаря состояниям возбуждения различные усвоенные нами уроки более глубоко записываются в каналах нашего мозга. Так, например, поездка лучше запоминается, когда вы нервничаете, пытаясь отыскать незнакомое место, а не когда вы следуете привычным маршрутом на автопилоте.
Источник: knife.media