Хлоропласты водорослей: строение и разнообразие
МЕНЮ
Главная страница
Поиск
Регистрация на сайте
Помощь проекту
Архив новостей
ТЕМЫ
Новости ИИ
Городские сумасшедшие
ИИ в медицине
ИИ проекты
Искусственные нейросети
Искусственный интеллект
Слежка за людьми
Угроза ИИ
ИИ теория
Компьютерные науки
Машинное обуч. (Ошибки)
Машинное обучение
Машинный перевод
Нейронные сети начинающим
Психология ИИ
Реализация ИИ
Реализация нейросетей
Создание беспилотных авто
Трезво про ИИ
Философия ИИ
Генетические алгоритмы
Капсульные нейросети
Основы нейронных сетей
Промпты. Генеративные запросы
Распознавание лиц
Распознавание образов
Распознавание речи
Творчество ИИ
Техническое зрение
Чат-боты
Авторизация
2020-06-30 23:15
Автор — Екатерина Рюмина
Под редакцией администрации группы
В 1905 г. Константин Мережковский (брат писателя Дмитрия Мережковского) выдвинул теорию симбиогенеза, в которой утверждал, что хлоропласты — это симбионты клеток. Его вывод был основан на уже тогда имеющихся данных о том, что хлоропласты никогда не синтезируются de novo, но воспроизводятся делением, которое может быть не синхронизировано с делением клетки.
Научное сообщество не приняло смелую теорию. Лишь в конце 1960-х она была переоткрыта и дополнена американским биологом Линн Маргулис. Сейчас же симбиотическое происхождение хлоропластов и митохондрий не вызывает сомнений. Более того, известно, что эндосимбиоз (и первичный, и вторичный) происходил не один раз. И это дает возможность проследить эволюционный путь хлоропластов — от бактерий до органелл самых разных групп организмов.
Итак, в этой статье речь пойдет об эволюции хлоропластов.
Так как хлоропласты — потомки бактерий, в них осталась бактериальная ДНК. Однако они стали органеллами, а это означает, что произошли три события. Первое — перенос генов из хлоропласта в ядро (это постепенный процесс, поэтому по числу перенесенных генов можно судить о давности эндосимбиоза). Второе — налаживание транспорта белков, синтезируемых теперь вне хлоропластов, в хлоропласты. И третье — синхронизация деления хлоропластов и клетки.
Непосредственно явление, при котором организм превращает другой свободноживущий организм в свою органеллу, называется эндосимбиоз. При этом первичным эндосимбиозом называется поглощение именно бактерии одноклеточным организмом. Сложно сказать, археей или эукариотом, потому что эти события тесно связаны с появлением ядра и, следовательно, эукариот, а значит, что было раньше, что позже — спорный вопрос. А вторичным эндосимбиозом называется «поглощение» одноклеточного эукариотного организма со своим хлоропластом другим одноклеточным эукариотным организмом.
Cyanophyceae (цианобактерии), или С чего все началось

Итак, началось все с первичного эндосимбиоза. Так кого же поглотил этот одноклеточный, еще не имевший пластид организм? Цианобактерию. Поэтому, чтобы понять устройство пластид эукариот, необходимо изучить строение первых цианобактерий.
Цианобактерии, или синезеленые водоросли, — это грамотрицательные бактерии, из чего следует, что у них две мембраны, внешняя и внутренняя, между которыми находится клеточная стенка. Казалось бы, это означает, что первичный хлоропласт должен был получиться с тремя мембранами (две от цианобактерии, одна от пищеварительной вакуоли хозяина). Но это не так. Скорее всего, была утеряна мембрана, соответствующая пищеварительной вакуоли. Например, она могла просто распасться из-за того, что симбионты внутри нее росли и размножались, а сама она — нет. Всякий факт есть факт: в результате первичного эндосимбиоза получились двумембранные пластиды, причем, скорее всего,обе они — бактериальные (об этом свидетельствует в том числе хлоропласт Glaucocystophyta, в котором между двумя мембранами сохранился слой муреина — вещества, из которого состоит клеточная стенка бактерий).
Цианобактерии — единственные оксигенные фототрофы среди бактерий. Использование воды в качестве донора электронов, а значит выделение кислорода стало возможным благодаря приобретению еще одной фотосистемы — фотосистемы 2 (далее — PS II). И эти две фотосистемы (PS I и PS II) с вытекающей Z-схемой фотосинтеза есть у всех фотосинтезирующих эукариот.
Из пигментов у разных групп цианобактерий найдены все хлорофиллы (a, b, c, d, f), а также каротиноиды и фикобилины, образующие фикобилисомы, — светособирающие комплексы на PS II. На рисунке 1 представлена типичная фикобилисома красных водорослей (вместе с электронотранспортной цепью фотосинтеза), но следует понимать, что у цианобактерий она немного другая (иначе цианобактерии тоже были бы красного цвета).

Где же происходит фотосинтез? PS I и PS II расположены в мембранных пузырьках, которые отшнуровываются от цитоплазматической мембраны, — тилакоидах (известен только один представитель, у которого нет тилакоидов, а фотосинтетический аппарат расположен непосредственно в плазмалемме — Gloeobacter violaceus).
А потом произошел первичный эндосимбиоз. Происходил он как минимум два раза: первый случай привел к образованию красных и зеленых водорослей (и глаукофитовых), которые потом дали пластиды всем остальным фотосинтезирующим группам. Второй же случай произошел с представителем надцарстваRhizaria — Paulinella chromatoohora.
Хлоропласт
Прежде чем обсуждать различия в строении хлоропластов у разных групп, рассмотрим общее строение хлоропласта на примере хлоропласта зеленого растения (см рис. 2). Итак, из чего он состоит?

- Внутреннее пространство хлоропласта называется строма, внутреннее пространство тилакоидов — люмен.
- Тилакоиды — структурная фотосинтезирующая единица хлоропласта. Это плоский мешок, окруженный мембраной, в которой находятся пигменты фотосинтеза. Тилакоиды могут быть одиночными (как у красных водорослей) или образовывать ламеллы. Стопки тилакоидов, соединенные ламеллами между собой, называются гранами.
- Пиреноид — уплотненная область стромы хлоропласта, в которой содержится фермент Рубиско (RUBISCO). Этот фермент фиксирует углекислый газ (СО2). Пиреноид может быть пронизан тилакоидами (как у зеленых и харовых водорослей) или нет (как у бурых водорослей). Наличие пиреноида — признак не слишком постоянный для отделов, поэтому далее рассматриваться не будет.
- Нуклеоморф — может сохраняться, если хлоропласт получен в результате вторичного или более эндосимбиоза (например, у гаптофитовых и криптофитовых).
- Хлоропластная кольцевая ДНК — свидетельство прокариотного происхождения хлоропластов. У разных групп редуцирована в разной степени (больше всего — у динофит). Она может храниться: в виде кольца (как у охрофитовых), в виде маленьких шариков (как у зеленых и харовых) и т. д.
- Рибосомы — опять же, прокариотное наследие.
- Глазок (стигма) — органелла, участвующая в фоторецепции. У некоторых это ряд пигментных глобул, расположенных в хлоропласте (например, зеленые водоросли) или цитоплазме (эвглены), у других — куда более сложная структура. Так как глазок нужен для фототаксиса, наблюдается корреляция между наличием жгутика и глазка. К примеру, глазков не бывает у красных, синезеленых и диатомовых водорослей, у которых нет и жгутиков.
Пигменты фотосинтеза — также немаловажный признак. Их делят на три группы:
• хлорофиллы (a, b, c...);
• каротиноиды: каротины (альфа и бета) и ксантофилы (зеаксантин, фукоксантин, перидинин и т. д.);
• фикобилипротеины — в типичном случае, это фикоэритрин, фикоцианин и аллофикоцианин, которые образуют фикобилисому.
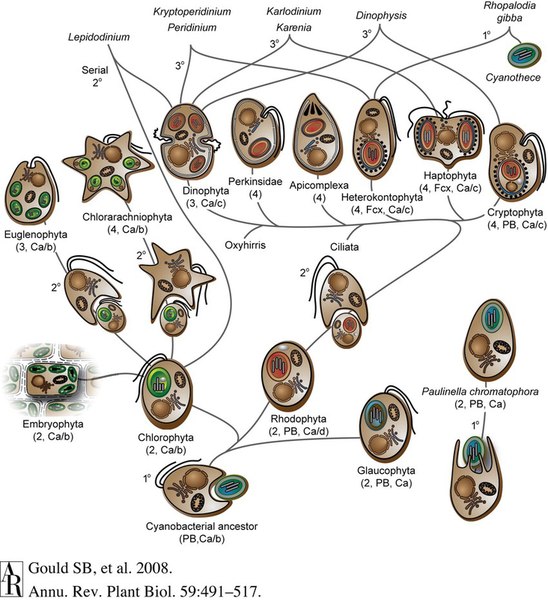
Как же происходила эволюция хлоропластов? И чем они различаются? На рисунке 3 представлена схема эволюции. В скобках указаны: число мембран, окружающих хлоропласт, вспомогательные пигменты фотосинтеза (PB — фикобилины, Fcx — фукоксантин) и присутствующие хлорофиллы. Цифрой у стрелочки обозначен порядок эндосимбиоза — первичный, вторичный, третичный.

Начнем с ветви красных водорослей — их самих и тех, кто получил хлоропласт, съев красную водоросль.
Rhodophyta (красные водоросли)

Как уже говорилось, красные водоросли получили свои пластиды в результате первичного эндосимбиоза (следовательно, хлоропласты окружены двумя мембранами). Поэтому у хлоропластов красных водорослей есть схожие с цианобактериями черты (помимо очевидных — таких как наличие ДНК и т. д.):
- Фикобилисомы. Здесь они как раз представлены в своем наиболее классическом виде (см рис. 1), то есть состоят из трех типов пигментов: фикоэритрина (внешний слой) красного цвета, фикоцианина и аллофикоцианина голубого цвета. У пресноводных представителей (например, у порядка Batrachospermales) нет фикоэритрина, из-за чего они имеют сине-зеленую окраску.
- Одиночные тилакоиды. Одиночные они из-за фикобилисом, которые сидят на мембране и не дают тилакоидам слипаться.
- Отсутствие глазка.
В таблице 1 сведена основная информация о хлоропластах красных водорослей. Следует, однако, понимать, что в ней (как и в последующих таблицах) разобраны типовые случаи и реального разнообразия они не покрывают.


В результате вторичного эндосимбиоза с красными водорослями свои хлоропласты получили группа SAR (Stramenopila, Alveolata, Rhizaria — включающая в себя динофитовые и охрофитовые водоросли, а также споровиков — см рис. 3), а также Haptophyta и Cryptorhyta. Последняя группа крайне важна, как переходная стадия между красными водорослями и SAR: уже нет фикобилисом, но есть хлорофилл с. У криптофит еще есть фикобилины (как у красных), но нет аллофикоцианина, и поэтому нет фикобилисом. При этом у них уже есть хлорофилл с (как у SAR), который понадобился как раз в связи с отсутствием фикобилисом. Родство этой группы с красными водорослями также подтверждается нуклеоморфом (остаточным ядром), доставшимся от оных.
SAR — Alveolata — отдел Dinophyta (динофиты, они же динофлагелляты)

Предок всей группы был фотосинтезирующим организмом, который получил хлоропласт, съев красную водоросль. И его хлоропласт окружен тремя мембранами. Но ведь в результате вторичного эндосимбиоза должно было получиться четыре мембраны: две от хлоропласта красной водоросли, одна — плазмалемма красной водоросли, и еще одна — пищеварительная вакуоль динофитовой водоросли. Так какая из мембран отсутствует и почему? Одно из возможных объяснений предполагает изначальное отсутствие мембраны, соответствующей плазмалемме красной водоросли. Базируется это объяснение на существовании у динофлагеллят мизоцитоза — способа питания, при котором хищником всасывается содержимое съедаемой клетки (без ее внешней мембраны). Способ этот удобен для динофит, у которых оболочка устроена очень сложно, — она представляет собой трехмембранную пелликулу. Однако в этом случае не очень понятно, как съеденная клетка без своей мембраны могла делиться (то есть как мог делиться получившийся трехмембранный хлоропласт). Другое объяснение предполагает изначальное наличие всех четырех мембран и последующую утерю все той же мембраны красной водоросли. Так что три мембраны динофит трактуются как две мембраны хлоропласта красной водоросли и одна мембрана динофитовой водоросли.
У динофит есть одна пренеприятнейшая особенность. Они часто теряли свои хлоропласты, а потом обзаводились новыми. И донором этих вторичных хлоропластов могло быть почти все: и диатомовые водоросли, и зеленые, и криптофиты и т. д. (на рис. 3 это показано стрелочками, соединяющими динофит с другими водорослями). Эти хлоропласты могут быть очень разными: в них могут оставаться ядро и митохондрии съеденного (в случае съеденной диатомовой водоросли) или же может перевариваться все, кроме двух мембран самого первого хлоропласта (как в случае со съеденными криптомонадами). А первичный хлоропласт к тому же может превращаться в глазок.
Из необычных пигментов присутствует перидинин, который вместе с хлорофиллом с и белками образует светособирающий комплекс (то есть выполняет функцию отсутствующих у SAR фикобилисом).
Также динофлагелляты — единственные эукариоты, у которых есть RUBISCO II (у всех остальных — I). Он отличается от RUBISCO I тем, что состоит всего из двух субъединиц (а не из восьми больших и 8 малых) и кодируется в ядерном геноме. Последнее свидетельствует о том, что у динофит сильно редуцирован хлоропластный геном: он содержит всего несколько десятков генов. Причем ДНК может быть разбита на отдельные кольцевые фрагменты.
Строение глазка также отличается большим разнообразием: это может быть как ряд глобул в цитоплазме или хлоропласте, так и более сложная структура, окруженная мембранами (как было сказано выше, такой глазок, видимо, получается из старого хлоропласта).

Помимо динофитовых водорослей в группе Alveolata есть еще одна группа, имеющая хлоропласты (пусть и редуцированные). Речь идет о споровиках (Apicomplexa) — вам наверняка знакомы хотя бы такие представители этой группы, как малярийные плазмодии. Для него, как и для некоторых других споровиков, характерно наличие апикопласта — бывшего хлоропласта. Он окружен 4 мембранами (что логично при вторичном эндосимбиозе), делится при делении клетки, не синтезируется de novo и т. д. Функция фотосинтеза им утрачена, однако он может быть задействован (у тех же малярийных плазмодиев) в синтезе веществ, привлекающих комаров.

SAR — Stramenopila — отдел Ochrophyta (охрофитовые водоросли)

Это очень большой отдел, включающий диатомовые, бурые, желтозеленые, золотистые и прочие водоросли. Скорее всего, вам больше всего знакомы бурые водоросли — ламинария, фукус и другие. Чем же сходны хлоропласты всех этих групп водорослей?
Все они окружены четырьмя мембранами, причем внешняя сливается с мембраной ядра (как и большинства любых водорослей с четырехмембранными хлоропластами). Из хлорофиллов присутствуют хлорофилл а и с, причем только у одного класса (диатомовые водоросли) присутствуют все три формы хлорофилла с (с1, с2, с3), у остальных же, как правило, есть только хлорофиллы с1 и с2. Также для этой группы характерен дополнительный пигмент — фукоксантин, который, однако, тоже есть не у всех (он отсутствует у желтозеленых и некоторых других водорослей, имеющих вошериаксантин). Кроме того, присутствуют каротиноиды. ДНК у большинства представителей собрана в кольцо, которое находится под опоясывающей ламеллой — еще одной характерной чертой.
Глазка нет у диатомовых водорослей (у которых нет жгутиков), а у бурых водорослей есть только у жгутиковых стадий (зооспор и гамет).


На этом рассмотрение групп, получивших хлоропласт от красных водорослей, заканчивается. И мы переходим к группе, получившей его от зеленых водорослей. И опять-таки логично начать с самих зеленых водорослей.
Viridiplantae — зеленые растения

Вполне вероятно, что вы видите это название первый раз в жизни. Что же это? Так называется группа, объединяющая зеленые и харовые водоросли, а также высшие растения. И одним из признаков, объединяющим эти группы, является как раз строение хлоропластов и набор пигментов.
Итак, вся это группа получила свои хлоропласты в результате первичного эндосимбиоза. Из чего логично вытекает их двухмембранность. У этой группы нет фикобилисом, что позволило собирать тилакоиды в граны. Имеются хлорофиллы а и b и дополнительные пигменты — каротиноиды. Они заслуживают отдельной статьи, однако некоторые стоит назвать: это бета-каротин (лидер по его процентному соотношению — Dunaliella), астаксантин (лидер — Haematococcus) и лютеин. Они могут выполнять самые разные функции: служить светособирающим комплексом или же в качестве антиоксиданта, обуславливать реакцию на температурный и осмотический шок, предохранять от избытка света. Кольцевые молекулы ДНК в хлоропласте собраны в шарики — нуклеоиды — и распределены по всему хлоропласту.
Для этой группы характерно хранение запасного вещества (крахмала) в строме хлоропласта (что больше несвойственно никому: остальные водоросли либо запасают не крахмал, либо откладывают его вне хлоропластов).


Excavata — отдел Euglenophyta (или Euglenozoa)

Нефотосинтезирующий предок этой группы получил хлоропласт в результате вторичного эндосимбиоза: он съел зеленую водоросль. Это объясняет набор хлорофиллов — a и b. Однако эта группа примечательна тем, что далеко не все ее представители имеют хлоропласты (есть как первично бесхлоропластные, так и вторично — утратившие хлоропласты). А все фотосинтезирующие являются миксотрофами. Поэтому представителей этого отдела рассматривают и зоологи, и альгологи, называя его соответственно своей номенклатуре (откуда и берутся два варианта: Euglenophyta, если вы альголог, и Euglenozoa, если вы зоолог).
Наличие всего трех мембран опять-таки поднимает все те же вопросы, что и в случае с динофитовыми водорослямми. Объяснения тут все те же. Покров эвглен — пелликула, которая состоит из мембраны, микротрубочек и белковых пластин. Этот сложный покров опять-таки объясняет наличие у эвглен мизоцитоза. Который в свою очередь объясняет (или нет) наличие всего трех мембран. Всяко три мембраны здесь все так же трактуются как две мембраны от съеденной водоросли и одна от хозяина.

В целом хлоропласт эвгленовых напоминает хлоропласт динофитовых, приведенный на рисунке 5.
• Ботаника в четырех томах, том 1 и 2 «Воросли и грибы» Г. А. Белякова, Ю. Т. Дьяков, К. Л. Тарасов / М.: Издательский центр «Академия», 2006
• Ботаника. Курс альгологии и микологии. Под редакцией Ю. Т. Дьякова / М.: Издательство МГУ, 2007
• Лекции Г. А. Беляковой по альгологии
•Plastid Evolution, Sven B. Gould, Ross F. Waller, and Geoffrey I. McFadden, 2008
• The endosymbiotic origin, diversification and fate of plastids, Patrick J. Keeling, 2010
• The Manganese-calcium oxide cluster of Photosystem II and its assimilation by the Cyanobacteria, James D. Johnson, M.S., Alumnus, Department of Chemistry, 2006
• Symbiogenesis: Mechanisms, Evolutionary Consequences, and Systematic Implications Thomas Cavalier-Smith, 2013
Телеграм: t.me/ainewsline
Источник: m.vk.com