Дарвинизм и палеоботаника: факты и гипотезы
МЕНЮ
Главная страница
Поиск
Регистрация на сайте
Помощь проекту
Архив новостей
ТЕМЫ
Новости ИИ
Городские сумасшедшие
ИИ в медицине
ИИ проекты
Искусственные нейросети
Искусственный интеллект
Слежка за людьми
Угроза ИИ
ИИ теория
Компьютерные науки
Машинное обуч. (Ошибки)
Машинное обучение
Машинный перевод
Нейронные сети начинающим
Психология ИИ
Реализация ИИ
Реализация нейросетей
Создание беспилотных авто
Трезво про ИИ
Философия ИИ
Генетические алгоритмы
Капсульные нейросети
Основы нейронных сетей
Промпты. Генеративные запросы
Распознавание лиц
Распознавание образов
Распознавание речи
Творчество ИИ
Техническое зрение
Чат-боты
Авторизация
2020-05-23 11:07

Палеоботаник Сергей Наугольных о коэволюции, развитии растений пермского периода и особенностях голосеменных растений
Крыло таракана. Жилки на крыле повторяют жилкование листьев птеридоспермов, встречающихся в тех же пермских отложениях. Печорское Приуралье
Если посмотреть на литературу по различным аспектам эволюционной теории, легко увидеть, что и среди профессиональных биологов, и среди людей, интересующихся естествознанием, нередки скептические, а то и едко-иронические высказывания об отдельных положениях дарвинизма и об эволюционной теории (теориях) в целом. Конечно, среди скептиков и опровергателей есть те, кто просто хочет самоутвердиться, противопоставив себя большинству коллег, и таким образом примкнуть к некоей «интеллектуальной богеме». Таких можно не принимать во внимание, но этим не устранятся некоторые из затруднений в дарвинизме как в любой развивающейся теории. Дальнейшее развитие дарвинизма и его подтверждение, безусловно, требуют фактического материала. Необходимыми подтверждениями и даже доказательствами изобилует геологическая летопись, а значит, и палеонтология. Поэтому палеонтологические данные (а может быть, они особенно) наравне с результатами современных биологических исследований необходимо вовлекать в дискуссии об эволюции органического мира.
Здесь я попытаюсь показать, что может дать палеоботаническая фактология для иллюстрации некоторых из наиболее важных положений дарвинизма.
Естественный отбор
Находки ископаемых остатков удивительных и необычных растений в земных слоях в свое время сыграли очень большую роль в развитии общественного сознания. Люди убедились в том, что когда-то жизнь на Земле была совершенно иной. Не менее важной была и кажущаяся теперь тривиальной изменчивость разных органов ископаемых растений, принадлежавших одному виду. Кстати будет заметить, что размах этой изменчивости в геологическом прошлом подчас существенно превышал нормы, свойственные современным растениям. А изменчивость как таковая, будучи одним из «трех китов» дарвинизма, поставляет материал для естественного отбора.
На палеоботаническом материале также удается выстроить филогенетические ряды от предка до потомка. И они красноречиво и однозначно подтверждают наследуемость признаков. А это еще одна движущая сила эволюции. Надо сказать, что во времена Дарвина однозначность наследуемости была далеко не так очевидна. Но можно ли на палеоботаническом материале проиллюстрировать работу естественного отбора?Разумеется, нельзя непосредственно увидеть действие естественного отбора, располагая лишь отпечатками листьев или других органов ископаемых растений. Однако можно заметить его результаты и уже потом строить гипотезы о действовавших в том или ином случае факторах отбора и эволюционных последствиях или морфологических решениях, к которым он мог привести. Для построения таких гипотез необходимо сопоставить филогенетические и морфогенетические последовательности в разных группах ископаемых растений с характером изменения реконструированных палеогеографических и палеоклиматических условий, в которых растения существовали.
Обычно при анализе палеоботанического материала трудно установить какую-нибудь эволюционную тенденцию или морфогенетический тренд, так как многочисленные наблюдения разнородны. Поэтому здесь требуется один и тот же методический прием. Я пошел именно этим путем, когда составлял схемы морфогенетических преобразований цикадофитов, гинкговых, пельтаспермовых, користоспермовых и кейтониевых. Для каждого временного уровня был взят наиболее типичный представитель (или два) данной филетической ветви, интерпретация которого наименее противоречива и для которого вы сока степень вероятности прижизненного сочетания разных органов. Это приближает статус анализируемого таксона (рода, вида) к естественному. К сожалению, не для всех анализируемых растений такое сочетание было установлено, однако с этим пришлось смириться. Увы, такова специфика палеоботанического материала. Далее отобранные для анализа таксоны выстраивались в последовательность, соответствующую геологическому возрасту или времени существования этих растений.
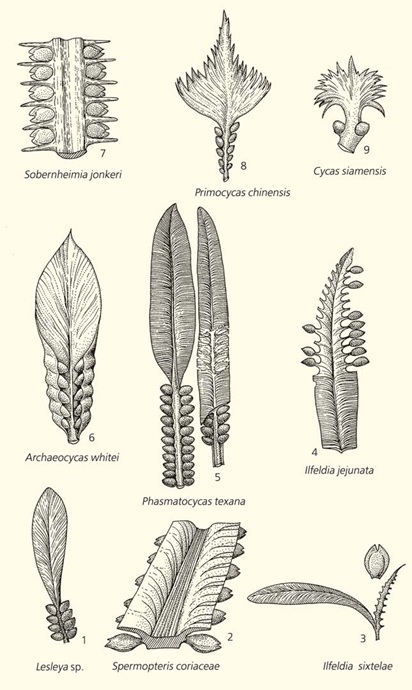
// Направление эволюции генеративных органов цикадовых. Видно, что у растений каменноугольного периода (1–4) семязачатки были множественными и располагались на рахисах или по краям листоподобных семеносных органов. В течение перми (5–8) у цикадовых постепенн
Начнем с цикадофитов. Древнейшие остатки растений, с той или иной степенью уверенности относимые к цикадовым (класс Cycadopsida), известны из верхнекаменноугольных отложений. Практически у всех позднепалеозойских цикадофитов, независимо от детальности интерпретации растительных остатков, семеносные органы представляли собой уплощенные листовые пластинки, по краям которых располагались семязачатки. У некоторых форм они прикреплялись к средним жилкам листьев, или осям, рахисам. Некоторые из ранних цикадофитов были еще настолько примитивными, что их можно бы отнести и к медуллозовым, или тригонокарповым, птеридоспермам, у которых перистые листья были немодифицированными, а семязачатки располагались на рахисах.
У пермских форм листовая пластинка семеносных органов постепенно редуцировалась, а число продуцируемых семян сократилось. У некоторых видов пластинка (в соответствии с ботаническими традициями ее можно назвать мегаспорофиллом) не исчезла совсем — ее края подвернулись и прикрыли семязачатки, как, например, у Archaeocycas whitei. Я думаю, эта тенденция далеко не случайна. В низких широтах Земли, где в основном и произрастали примитивные цикадофиты, в течение всей перми климат постепенно становился все более и более засушливым.[1 ] Позднекаменноугольные цикадофиты имели широкие, хорошо развитые мегаспорофиллы, приспособленные к испарению большого количества влаги в условиях влажных лесов. В засушливом климате пермского периода эти приспособления оказались не только ненужными, они представляли собой фатальный морфологический груз. Защита семязачатков подвернутыми краями мегаспорофиллов также могла быть селективной реакцией на иссушение климата. Она и была закреплена у части цикадофитов естественным отбором, а менее приспособленные формы вымерли в течение раннепермской эпохи.
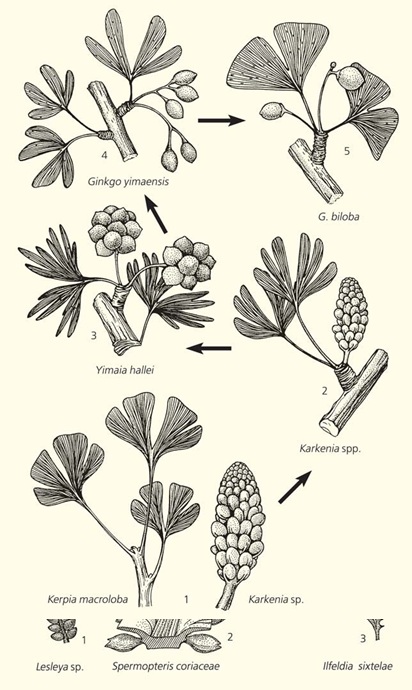
// Эволюционная тенденция в морфогенезе семеносных органов гинкгофитов. Эта тенденция выразилась в неуклонном снижении количества семязачатков от ранней перми (1) и до современности (5). 1 — S.V.Naugolnykh; 2 — обобщенная реконструкция, S.Archangelsky, 1965,
Очень сходная тенденция, правда, выразившаяся в уменьшении количества продуцируемых семязачатков, проявляется в морфогенезе семеносных органов гинкгофитов (порядок Ginkgoales). Роль гинкговых и цикадофитов в эволюции растительности на рубеже палеозоя и мезозоя очень похожа. Те и другие появились в позднем карбоне, а в перми уже существовал целый ряд форм, хотя и весьма малочисленных. Доминировали тогда древние группы голосеменных — кордаиты, диктиоптеридиевые (глоссоптериды), войновскиевые и палеозойские семейства хвойных.
После экосистемной перестройки на рубеже перми и триаса подавляющее большинство групп растений, типичных для палеозоя, вымирает. Начинается постепенный рост разнообразия и количественного участия в растительном покрове гинкговых и цикадовых. И уже с позднего триаса они становятся одними из определяющих элементов мезозойской растительности.
Уменьшение количества продуцируемых семян у гинкговых можно объяснить повышением эффективности репродуктивных механизмов, жизненно необходимых в условиях неуклонной аридизации климата и обострившейся в связи с ней борьбой за существование. Типичные для палеозоя кордаиты, войновскиевые и глоссоптериды вымерли в ходе этой борьбы под действием жестокого естественного отбора на фоне прогрессирующего иссушения климата.
Постепенное вымирание гинкговых в течение кайнозоя совпадает по времени с исчезновением тех млекопитающих, которые поедали их семена и тем способствовали распространению этих растений. У прошедших через пищеварительный тракт семян твердая оболочка семян размягчалась, и облегчалось прорастание заростков, которые к тому же получали дополнительные органические вещества и влагу, необходимые для первой фазы онтогенетического развития. Тот, кто сталкивался со зрелыми семенами современного гинкго, наверняка запомнил их странный, острый и, в общем-то, неприятный запах. Его источают мягкий воротничок и внешние покровы семени, окрашенные в желто-оранжевый цвет. И яркая окраска, и сильный запах, подобно феромонам, притягивали животных, питавшихся семенами гинкговых. Раньше, в мезозое, это были, конечно же, не млекопитающие, а разнообразные наземные и летающие рептилии. Их вымирание на рубеже мела и палеогена также могло способствовать постепенному снижению количества гинкговых в палеофитоценозах.
У наиболее древних, палеозойских, гинкгофитов настоящего воротничка еще не было, однако у ранннеюрских он уже хорошо развит. Происхождение воротничка до сих пор остается не вполне определенным. Полагаю, что он мог образоваться за счет совмещения признаков аберрантных воронковидных листьев с другой аберрацией — заложением семязачатка на краю листовой пластинки.[2 ] Произойти это должно было как раз в конце перми — начале триаса. Так может быть, появление воротничка, привлекавшего внимание некрупных растительноядных рептилий, и повысило эффективность репродукции гинкговых на рубеже палеозоя и мезозоя? Именно эта мутация могла быть подхвачена естественным отбором и закреплена в последующих поколениях гинкгофитов.

// Moresnetia sp., одно из наиболее примитивных голосеменных растений (реконструкция и отпечаток в породе). У этого раннекаменноугольного птеридосперма семеносные органы представляли собой купулы (капсулы с открытой верхушкой), собранные в пучки на концах фе
Еще одна крупная филогенетическая ветвь тянется из палеозоя к современности, связывая на первый взгляд очень разрозненные формы. В основании ветви находятся весьма интересные и своеобразные голосеменные растения из семейства пельтаспермовых (Peltasperma ceae). В соответствии с традиционными представлениями, оно входит в состав одноименного порядка Peltaspermales, принадлежащего большому и разнообразному классу птеридоспермов, или иначе «семенных папоротников» (их еще называют Pteridospermopsida, Ly ginopteridopsida, Cycadofilicop sida).
Пельтаспермовых, конечно же, нельзя считать типичными птеридоспермами, ведь у них, в отличие от классических представителей этого класса, семеносные органы были уже в высокой степени специализированы — представляли собой уплощенные зонтиковидные образования. К нижней стороне этих семеносных дисков, или пельтоидов, прикреплялись семена. Такое строение характерно для типичных пельтаспермовых, собственно относящихся к роду Peltaspermum.
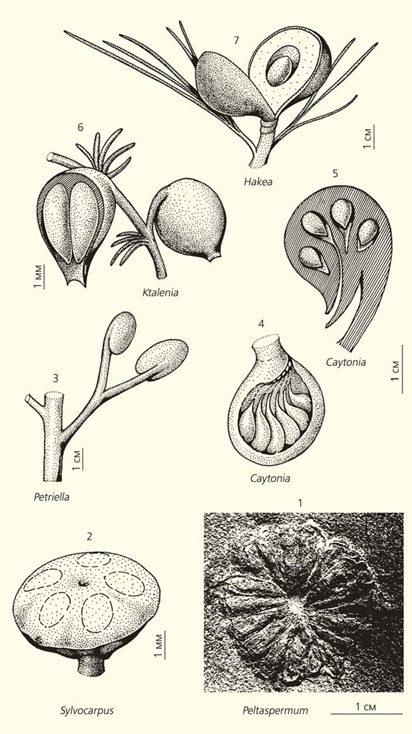
// Морфогенетический тренд в развитии семеносных органов. Уже в перми (1, 2) у пельтаспермовых (1–3) семеносные органы начали замыкаться, в триасе (3), юре (4, 5) и в мелу (6) процесс все более эффективной защиты созревающих семязачатков продолжался. В резул
Именно от них в конце раннепермской эпохи ответвилась новая группа родов, объединенных в другое семейство — ангаропельтидиевых (Angaropeltidaceae). Тогда оно состояло из двух родов — Sylvocarpus и Permoxylocarpus, отличавшихся строением женских репродуктивных органов: семеносные диски (какие были у пельтаспермовых) преобразовались в полузамкнутые (у Permoxylocarpus) или даже почти полностью замкнутые (у Sylvocarpus) семеносные капсулы. Это морфологическое изменение возникло за счет опускания краев диска и их смыкания вокруг ножки репродуктивного органа. Количество продуцируемых семян у пельтаспермовых и ангаропельтидиевых варьировало в весьма широких пределах, но и здесь улавливается не который эволюционный тренд: у раннепермского Peltaspermum retensorium оно достигало 25, у Permoxylocarpus trojanus — от 14 до 16, а у Sylvocarpus arma tus — всего пяти. Корреляция, намечающаяся между степенью замкнутости диска и количеством семян, может отражать повышенную эффективность защиты семязачатков во время их созревания. Чем лучше защита, тем меньше семян требуется для успешного размножения растения.
Последние достоверные пельтаспермовые вымерли в конце триасового периода. Но еще в его начале и подчас в тех же фитоценозах (особенно в Южном полушарии на материках Гондваны) уже существовали по соседству прогрессивные птеридоспермы — растения семейства користоспермовых (или, что-то же: умкомазиевых, дикроидиевых). Они, сохраняя многие черты пельтаспермовых, все же выходят на новый уровень организации. У некоторых користоспермовых (например, у растений родов Petriella и Ktalenia) семеносные органы практически полностью замкнуты. Этим они очень напоминают плодолистик покрытосеменных. У многих видов из родов Corystospermum и Ktalenia количество продуцируемых семян сокращается до двух.В юре и раннем мелу на смену користоспермовым приходят кейтониевые (семейство Cay toniaceae). По уровню организации они настолько приближаются к покрытосеменным, что непосредственных предков этой группы растений некоторые палеоботаники склонны искать именно среди кейтониевых. Конечно, у них еще не было цветка, но, как предполагается, их пыльца уже прорастала к семязачаткам через некое подобие рыльца, как у покрытосеменных.[3 ]
Сейчас нельзя однозначно утверждать, что пельтаспермовые — користоспермовые — кейтониевые — покрытосеменные связаны в единую филогенетическую цепочку. Однако здесь на лицо морфогенетический тренд, который отражает эволюционную тенденцию: она выражена во все более и более эффективной защите созревающих семя зачатков от неблагоприятных воздействий окружающей среды, послуживших в этом процессе самым активным фактором естественного отбора.
Разумеется, действие отбора в геологическом прошлом не ограничивалось уровнем таксонов, но распространялось и на эволюцию сообществ. И здесь отбор мог играть не только прогрессивную, созидательную роль, но и, напротив, иногда приводил к смене высокоорганизованных и сбалансированных сообществ на более примитивные. Один из самых показа тельных примеров в этом от ношении — изменение растительности в середине пермского периода в Приуралье, где на рубеже кунгурского и уфимского веков резко, можно сказать катастрофически, изменилась структура растительных сообществ. Разгар кризиса, его парадоксальная фаза, приходится на соликамское время — первую половину уфимского века. В Приуралье исчезает исключительно богатая видами кунгурская, или, как ее еще называют с легкой руки выдающегося российского палеоботаника М. Д. Залесского, бардинская, флора. Ее сменяет соликамская, или вячеславиевая, флора, так сономически очень бедная. Во многих местонахождениях она представлена единственным видом — древовидной вячеславией (Viatcheslavia vorcutensis), гетероспоровым плауновидным растением.
В конце раннепермской эпохи в Приуралье были и весьма высокоорганизованные растения, например разнообразные хвойные, птеридоспермы и гинкгофиты. Подавляющее большинство видов кунгурской флоры уже хорошо приспособлено к окружающей обстановке. Многие из них занимали строго определенные экологические ниши, о чем свидетельствуют различные морфологические и анатомические признаки — маркеры специфических адаптаций. Сами кунгурские палеофитоценозы были отчетливо структурированы и сложно организованы.
Соликамская растительность, напротив, была устроена довольно просто. Тафономические данные указывают: соликамские палеофитоценозы состояли из одного-двух катениальных звеньев, причем без отчетливой высотной структуры. Вячеславии, судя по тому, что их остатки встречаются в отложениях морского, лагунного и речного происхождения, могли произрастать в самых разных условиях, разумеется, если случались хотя бы кратковременные влажные сезоны, необходимые для размножения.
Чем же обусловлена эта весьма контрастная перестройка, столь глубоким образом отразившаяся на структуре расти тельных сообществ? В нижней части соликамских отложений во многих районах Северного, Среднего и Южного Приуралья есть несомненные признаки резкой аридизации климата: гипс и мощные прослои строматолитов. Это явное свидетельство ненормальной, то резко повышавшейся, то, наоборот, понижавшейся солености морского бассейна, в котором они образовались. Существовал он в Предуральском краевом прогибе и за крылся в конце раннепермской эпохи, после чего распался на отдельные лагуны.
Резкое иссушение климата в соликамское время во всем приуральском регионе и привело к вымиранию богатой видами кунгурской растительности. Значительная часть споровых и голосеменных растений не смогла приспособиться к резко изменившимся условиям. На смену этим растениям пришли менее специализированные и невзыскательные в экологическом отношении вячеславии, которые к тому же еще и неплохо переносили длительные засушливые периоды. Снова естественный отбор, но уже на уровне сообществ. Однако, в отличие от упомянутых таксономических примеров, он привел не к структурному прогрессу, а скорее к регрессу, правда временному.
Очень сходной, но уже в глобальном масштабе, была ситуация на рубеже перми и триаса. Тогда в роли пионеров, колонизовавших освободившиеся после кризиса пространства, выступили плевромейи. Будучи прямыми потомками приуральских вячеславий, они во многом напоминали предков морфологически. Это сходство, конечно же, тоже не было случайным. Аридизация климата на границе перми и триаса сделала необходимыми те же адаптации, что и во время соликамского кризиса в Приуралье.
Справедливости ради надо сказать, что примеры катастрофически быстрых изменений в растительном мире не позволяют согласиться с одной из наиболее широко распространенных максим Дарвина, что природа «не устраивает революций». Конечно же, устраивает, и не реже, чем они происходят в человеческом обществе, однако резкие изменения — «революции» — в биосфере протекают все по тем же универсальным эволюционным законам.
Прогрессивная эволюция
Перейдем от палеоботанических картин, иллюстрирующих механизмы и факторы эволюционного процесса, к их филогенетическому «овеществлению». Но прежде остановимся на прогрессивной эволюции и неразрывно с ней связанным положением классика дарвинизма А. Н. Северцова о ключевых ароморфозах.[4 ]
Анализируя пути развития растительного мира, можно видеть несколько узловых, ключевых этапов становления морфологических прототипов в различных группах высших растений. Само их появление до конца еще не ясно, хотя над этой проблемой исследователи бьются, пожалуй, со времени возникновения палеоботаники как науки. Очень многое сделано в последние десятилетия, и сейчас в общих чертах появление высших растений можно реконструировать.
Зарождение высших растений началось с анатомоморфологических приобретений: плотных покровов с устьицами (органами газообмена — «дыхания») и проводящей системы. Споры с трехлучевой щелью разверзания, характерные для многих споровых растений и вмещающие специализированные органы, видимо, появились чуть раньше, у произраставших на сублиторали и литорали водорослей. Экологически они близки современным бурым водорослям — фукусам и аскофиллюмам.
Становление прототипа высших растений завершилось в силуре, и далее началось активное совершенствование примитивных форм, приспособление к условиям среды, приведшее к вспышке биоразнообразия. Его пик пришелся на ранний и средний девон. Крупные морфологические преобразования (появление эпидермиса и других специализированных тканей, кутикулы, устьиц и трахеид) позволили споровым растениям выйти на качественно новый эволюционный уровень и освоить разнообразные околоводные пространства в середине палеозоя.
В конце девонского периода в растительном мире возникает новая репродуктивная система, связанная с образованием семян. Благодаря этому растения обрели возможность предохранять зародыш нового организма, а за счет таких приспособлений для полета, как крылатки и хохолки на семенах, могли расселяться ветром на все новые и новые территории. Вслед за появлением первых семян примитивные голосеменные дали настоящую вспышку разнообразия в начале каменноугольного периода. Второе крупное приобретение — появление семян, т. е. органов воспроизведения и расселения, — открыло растениям пути для освоения «первичных пустынь» — удаленных от водоемов и водотоков возвышенных и водораздельных пространств.
Разумеется, далеко не все эволюционные процессы в растительном мире шли по схеме ароморфоз — идиоадаптация, т. е. по закону Северцова. Проще говоря, за крупными перестройка ми не всегда наступал период частных приспособлений. Сейчас установлено, притом на довольно обширных материалах из истории растений и животных, что некоторые морфогенетические тенденции возникали независимо в разных параллельно развивающихся группах. Таков, например, феномен «ангиоспермизации», постепенного выхода различных групп голосеменных на уровень развития покрытосеменных.
По мнению А. Г. Пономаренко, известного палеонтолога аналитика, «Новая группа не возникает в результате одного ключевого эволюционного преобразования и диверсифицируется в новой нише (или лицензии), а отдельные ее черты по являются у разных представителей групп предшественников, и именно эти новые свойства преобразуют мир, делая новую группу все более приспособленной, и в конце концов ее по явление становится неизбежным».[5 ]
С этой точки зрения, на определенном этапе эволюции сразу в нескольких группах голосеменных растений обозначилось направление, ведущее к прогрессивным преобразованиям. У продвинутых голосеменных это выразилось, прежде всего, в облекании семян все более и более замкнутыми оболочками — прообразами плодолистика покрытосеменных растений.
Существование общих тенденций в эволюции органического мира — не только «ангиоспермизации», но и других «заций» (атроподизации, т. е. приобретения признаков членистоногих разными исходными группами примитивных беспозвоночных; маммализации — появления признаков млекопитающих у нескольких групп зверообразных рептилий и т. д.) может быть понято как подтверждение ортогенеза, «направлен ной эволюции». А тогда отрицается роль дарвиновского естественного отбора, основанного на стохастических и вероятностных процессах.
Однако все эти «зации» можно объяснить и с позиций классического дарвинизма. Если организмы существуют в сходных условиях, причем не только абиотических, но и ценотических, то возникают и сохраняются сходные адаптации. Следовательно, эти общие тенденции есть не что иное, как такие адаптации — один из важных шагов в конкурентной борьбе растений за выживание.
Коэволюция
В «Происхождении видов» Дарвин пишет: «…Про растение, ежегодно производящее тысячу семян, из которых в среднем вызревает лишь одно, еще вернее можно сказать, что оно борется с растениями того же рода и других, уже покрывающими почву. Омела зависит от яблони и еще нескольких деревьев, но было бы натяжкой говорить о ее борьбе с ними потому только, что если слишком много этих паразитов вырастет на одном дереве, оно захиреет и погибнет. Но про несколько сеянок омелы, растущих на одной и той же ветви, можно совершенно верно сказать, что они ведут борьбу друг с другом. Так как омела рассеивается птицами, ее существование зависит от них, и, выражаясь метафорически, можно сказать, что она борется с другими растениями, приносящими плоды, тем, что привлекает птиц пожирать ее плоды и, таким образом, разносить ее семена. Во всех этих значениях, нечувствительно переходящих одно в другое, я … прибегаю к общему термину „Борьба за существование“».[6 ]
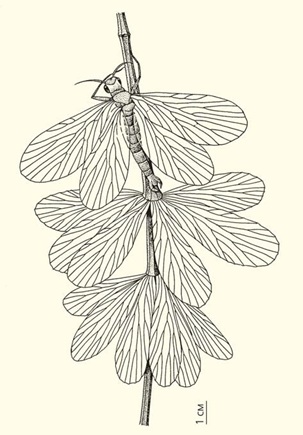
// Скорпионница (<em>Petromantis sylvaensis</em>) на клинолистнике (<em>Sphenophyllum biarmicum</em>). Сидя на побеге, петромантис маскируется под листовую мутовку, чтобы стать незаметным для пролетающих мимо хищных насекомых, например, гигантских стрекоз ме
Действительно, прогрессивная эволюция организмов и в особенности растений протекает в тесной связи с развитием их соседей и партнеров по биоценозам. Не знаю, можно ли точно подсчитать соотношение адаптаций организма к абиотическим факторам среды и приспособлений к сосуществованию с другими организмами одного и того же сообщества. Но совершенно очевидно, что приспособления к эффективному взаимодействию с другими компонентами биоценоза исключительно важны. Появляются такие симбиотические адаптации в геологической истории растений довольно рано. Уже в карбоне есть масса примеров коэволюции растений и животных (прежде всего членистоногих), их активного взаимодействия между собой. Очевидно, симбиотическая пара животное — растение возникла гораздо раньше.

// Крыло таракана. Жилки на крыле повторяют жилкование листьев птеридоспермов, встречающихся в тех же пермских отложениях. Печорское Приуралье.
В растительных сообществах конца палеозоя возникают разные симбиотические отношения между организмами. Пожалуй, хрестоматийный пример — мимикрия палеозойских тараканов: жилкование их крыльев очень похоже на жилкование перышек некоторых птеридоспермов. Мимикрия была свойственна также раннепермскому насекомому из группы скорпионниц — петромантису (Petromantis sylvaensis). Жилкование и очертания его крыльев и листьев сфенофиллума, или клинолистника (Sphenophyllum biarmicum), практически идентичны. Прекрасный пример творческой роли естественного отбора! Спрятавшись между листьями растения или затаившись на его побеге, насекомое становилось невидимым как для врагов, так и для потенциальной жертвы, что, естественно, резко повышало шансы на выживание и оставление потомства.
) того времени охотно прогрызали спорангии и семязачатки древних голосеменных и высасывали питательное содержимое. Разумеется, такой тип взаимодействия насекомых с растениями не мог не отразиться самым негативным образом на эффективности размножения последних, поскольку количество вызревающих семян неизбежно уменьшалось.

// Яйцекладка насекомого в желобке рахиса птеридосперма или папоротника. Еще один пример симбиотических отношений растений и насекомых. Нижнепермские отложения Среднего Приуралья, местонахождение Чекарда
Пищевые последовательности, связывающие воедино все компоненты экосистемы, разумеется, обусловливали относительную синхронность эволюционных преобразований и у продуцентов, и у консументов. Надо сказать, особую роль положения организма в пищевых цепочках для эволюционных преобразований неоднократно подчеркивал И. И. Шмальгаузен.[8 ]

// Микроперфорация на семеносной капсуле сылвокарпуса (<em>Sylvocarpus armatus</em>). Прокалывая толстую внешнюю кутикулу, насекомое высасывало питательное содержимое капсулы. Это пример трофической связи насекомых с растениями в раннепермское время. Диаметр
Изучая кутикулы упомянутого уже раннепермского сылвокарпуса, я обратил внимание на странные округлые дырочки перфорации, идущие по внешней поверхности.[9 ] Кутикулы оказались необычно толстыми, перфорации не пронизывали их насквозь, а по краям были видны травматические ткани. Видимо, семенами сылвокарпуса, заключенными внутри семеносной капсулы, питались какие-то насекомые со стилетом — приспособлением для прокалывания растительных тканей (такие стилеты известны у тех же палеодиктиоптер). Развитие мощной покровной кутикулы, предохранявшей капсулы и семена от повреждений насекомыми, было адаптивным ответом сылвокарпуса на их агрессию. И таких примеров в истории взаимоотношений растений и насекомых в истории Земли было немало.
* * *
В завершение вернусь к записке, переданной любознательным школьником. Так есть ли альтернатива дарвинизму в объяснении природы биологических (в том числе и палеоботанических) феноменов и процессов?
Если не принимать во внимание разнообразные теологические толкования, которые без труда могут объяснить вообще все что угодно, то, пожалуй, самой серьезной концепцией, которая конкурирует с дарвинизмом, можно считать теорию номогенеза Л. С. Берга.[10 ] Сходные представления в разные годы высказывались в той или иной форме разными биологами (например, А. Лима де Фариа [11 ]).
Номогенетические теории довольно красиво объясняют морфологическую структурированность организмов. Однако феномен «стрелы времени», прогрессивного развития биосферы Земли без привлечения естественного отбора как фактора, определяющего это развитие, они, на мой взгляд, объяснить не в состоянии. Именно поэтому дарвинизм в его современном понимании, опирающийся и на традиционные положения, и на популяционную генетику, остается основой основ современного естествознания и научной картины мира.
Телеграм: t.me/ainewsline
Источник: postnauka.ru