Серотонин: Краткий обзор мировосприятия
МЕНЮ
Главная страница
Поиск
Регистрация на сайте
Помощь проекту
Архив новостей
ТЕМЫ
Новости ИИ
Городские сумасшедшие
ИИ в медицине
ИИ проекты
Искусственные нейросети
Искусственный интеллект
Слежка за людьми
Угроза ИИ
ИИ теория
Компьютерные науки
Машинное обуч. (Ошибки)
Машинное обучение
Машинный перевод
Нейронные сети начинающим
Психология ИИ
Реализация ИИ
Реализация нейросетей
Создание беспилотных авто
Трезво про ИИ
Философия ИИ
Генетические алгоритмы
Капсульные нейросети
Основы нейронных сетей
Промпты. Генеративные запросы
Распознавание лиц
Распознавание образов
Распознавание речи
Творчество ИИ
Техническое зрение
Чат-боты
Авторизация
2018-10-15 13:34

Автор: Anonimch
Изображения: Антон Осипенко
Редакция: Deepest Depths, Алексей Дорохов, Михаил Гусев, Диана Мавлютова
Данная статья была опубликована на сайте «Medach», 03.04.2016.
Вот и пришло время написать о самом распиаренном и самом известном нейромедиаторе – серотонине. Самое интересное заключается в том, что о нём несколько десятилетий несут бред самые различные слои общества – начиная от домохозяек и гуманитариев, с их мифами о «гормоне счастья», заканчивая мамкиными психонавтами, которые скинулись однажды на марочку с ДОБом и теперь готовы каждому прохожему рассказывать о прелестях «изменённого состояния сознания».
Изначально считалось, что серотонин влияет только на тонус сосудов и гладких мышц, его прямое присутствие в ЦНС было обнаружено лишь в 1953 году Ирвином Пейджем и Бетти Твэрог, информацию о своём открытии они опубликовали в The American journal of physiology – «Serotonin content of some mammalian tissues and urine and a method for its determination.».
В каждой из таких ситуаций история поступает с наукой одинаково: любая недостаточно хорошо исследованная область начинает обрастать спекуляциями, либо действительно важные и серьёзные опыты и публикации теряются в общем шуме. Взрывы снарядов и свист пуль во время второй мировой войны заглушили опыт Альберта Хоффмана с ЛСД, а песни о всеобщей любви в 60е оказались более увлекательными, чем работы Шульгина и Николса. Думаю, что не стоит скрывать факт того, что 90% мировой «психоделической культуры» завязано на веществах, тем или иным образом влияющим на серотонинэргические нейроны в ЦНС.
Так что рассказ о серотонине стоит начать не с Хоффмана, Шульгина или Твэрог, а с краткого повествования об истории изменённых состояний сознания.
Повествование об изменённых состояниях сознания начнём с рассказа об эволюции мозга. В 2009 г. в «Троицком варианте» генетик Михаил Гельфанд опубликовал замечательную статью под названием «Шизофрения как последствие эволюции мозга» [1]. Главный вывод которой я позволю себе сформулировать более ёмко, чем это было в источнике – человек познакомился с безумием в момент становления человеком. При этом слово «момент» в контексте эволюционной нейробиологии можно смело принимать за несколько десятков тысяч лет. Учёные анализировали выборку генов-кандидатов, мутации в которых наиболее часто обнаруживаются у людей, больных шизофренией, и сравнили их с генами здоровых людей и генами наших ближайших родственников – шимпанзе и макак-резус. В итоге выяснилось, что большая часть генов-кандидатов влияла на уровень метаболизма нервных клеток и, соответственно, в случае человека являлась одной из наиболее быстро эволюционирующих групп генов. Так что шизофрению в контексте конкретно данного исследования можно считать «детской болезнью» в эволюционном пути человека.
В этот момент некоторые люди приобрели возможность с рождения воспринимать мир несколько иначе, чего не было у шимпанзе. Но человеку этого было мало – он требовал продолжения банкета! Тут как раз помогло другое эволюционное приобретение – фермент АДГ (алкогольдегидрогеназа), позволивший есть забродившие плоды и, чуть позже – со времени изобретения керамики, пить испортившийся фруктовый сок.
А что делать с похмелья? Конечно же! Искать всяческие корешки и пробовать – вдруг поможет?
Таким вот нехитрым образом древнее человечество открыло для себя первые лекарства, красители и яды. Среди случайно поглощённых растений с большой долей вероятности, т.к. прародина человечества была в Африке, был ибога. Этот небольшой кустарник — культовое растение для народности митсого, чей религиозный культ «бвити», возможно, является потомком самых древних практик направленного изменения сознания, живым реликтом психоделики. Интересно, что содержащийся в ибоге ибогаин и его метаболит – норибогаин – являются одновременно лигандами опиоидных и сертониновых (5-НТ3, ионотропный подтип) рецепторов [3].
Во многих древних культурах, где было распространено ритуальное использование растений, содержащих изменяющие сознание вещества, эти самые вещества зачастую воздействовали именно на серотонинэргическую нейротрансмиссию. Можно ещё вспомнить мифический напиток богов древней Индии – амриту, которая, согласно преданиям, готовилась из эфедры и грибов. Также, в качестве более подробно задокументированного примера, можно привести аяхуаску из Южной Америки. Интересно, что древние индейцы каким-то образом умудрились придумать такую смесь, которая содержит и ДМТ (диметилтриптамин, сильный серотонинэргический психоделик), и ИМАО (ингибиторы моноаминоксидазы) растительного происхождения. В нормальных условиях ДМТ неактивен при пероральном приёме, однако он активируется в смеси с иМАО, замедляющими его распад в ЖКТ.
На Руси наши предки также иногда ловили домотканые и лубяные приходы, прямо в печи деревенской выпеченные. Называлось это дело «пьяный хлеб» — то есть хлеб, выпеченный из муки злаковых растений, поражённых грибком. Интересно, что под этим понятием подразумевалось сразу два вида грибка — Fusarium graminearum, содержащий вомитоксин, и Claviceps purpurea (спорынья), содержащий производные эрготамина. И если отравления вомитоксином протекали относительно легко, то отравления спорыньёй даже получили своё название – эрготизм. При эрготизме, помимо острого психоза, нарушается тонус кровеносных сосудов. Это может привести к некрозу конечности или гангрене.
С наступлением нового времени и плавным переходом алхимии в химию появилась возможность выделять действующие вещества из растительных источников. Изначально любой экстракт содержал просто спиртовой раствор суммы алкалоидов — азотосодержащих веществ, которые, если говорить упрощённо, возможно было выделить стандартной кислотно-щелочной экстракцией. Затем люди научились выделять отдельные вещества из общего экстракта. Это дало возможность наблюдать за «чистым» фармакологическим действием отдельных химических веществ на организм.
В этом отношении серотонинэргическая система является середнячком: несмотря на то, что первые сообщения о «субстанции, влияющей на тонус мышц» относятся к XIX веку, серотонин в чистом виде был выделен лишь в 1935 году Витторио Эрспамером. В дальнейшем это вещество обнаруживалось во многих тканях и органах, в т.ч. и в нервной системе (см. выше). Однако, первые сведения о серотониновых рецепторах относятся лишь к 1957 году: Дж. Гаддум обнаружил, что способность серотонина сокращать гладкую мускулатуру можно блокировать ЛСД (диэтиламидом D-лизергиновой кислоты, в отечественной литературе также использовалось сокращение ДЛК), а морфином можно предотвратить возбуждение вегетативных ганглиев, наступающее при прямом воздействии серотонина. Таким образом, вплоть до 90х годов ХХ века серотониновые рецепторы были разделены лишь на 2 класса – Д- и М-рецепторы. Сейчас, благодаря методам молекулярной биологии, открыто 7 классов серотониновых рецепторов, причём многие из них имеют несколько подклассов (подтипов).
Также стоит упомянуть и о том, что серотонин – это не только нейротропный гормон, чьи функции ограничены лишь пределами нервной системы. Например, он участвует в процессах свёртывания крови и пищеварения. Более того, в 2003 году был введён термин «серотонилирование» (serotonylation)[4], описывающий прямое действие серотонина на внутриклеточные белки. Этот процесс был описан относительно триггерного механизма, определяющего высвобождение сигнальных веществ из бета-клеток поджелудочной железы и активацию тромбоцитов.
Метаболизм серотонина
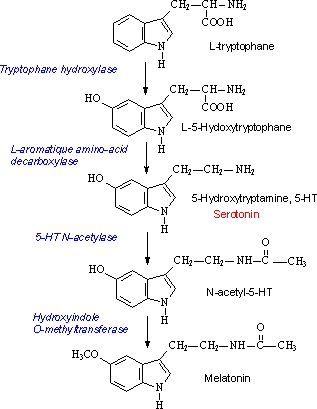
Изначально триптофан под действием фермента триптофангидроксилазы получает ОН-группу по 5-положению в основном индольном фрагменте, превращаясь в 5-гидрокситриптофан (5-HTP в англоязычных источниках). Затем 5-НТР путём отщепления –СООН группы декарбоксилазой ароматических аминокислот превращается в собственно серотонин.
Дальнейший метаболизм серотонина разветвлён и зависит от «ферментного набора» ткани и органа, где соответственно протекает [5].

Однако, наибольшую значимость имеет путь, ведущий к мелатонину. Фермент N-ацетилтрансфераза превращает серотонин в N-ацетилсеротонин, а затем 5-гидроксииндол-О-метилтрансфераза превращает ОН-группу в CH3O-, осуществляя таким образом финальную стадию синтеза мелатонина (N-ацетил-5-гидрокситриптамина).
Также интересен распад (катаболизм) серотонина. В нём задействован фермент моноаминоксиадаза (МАО), о котором упоминалось ранее. Он превращает серотонин в 5-гидроксииндолилацетальдегид (5-HILA на схеме). Затем этот альдегид может быть либо восстановлен альдегидредуктазой (ALDR) до 5-гидрокситриптофола, либо путём взаимодействия с альдегиддегидрогеназой 2 (ALDH2) превратится в 5-гидроксииндолилуксусную кислоту (5-HIAA).
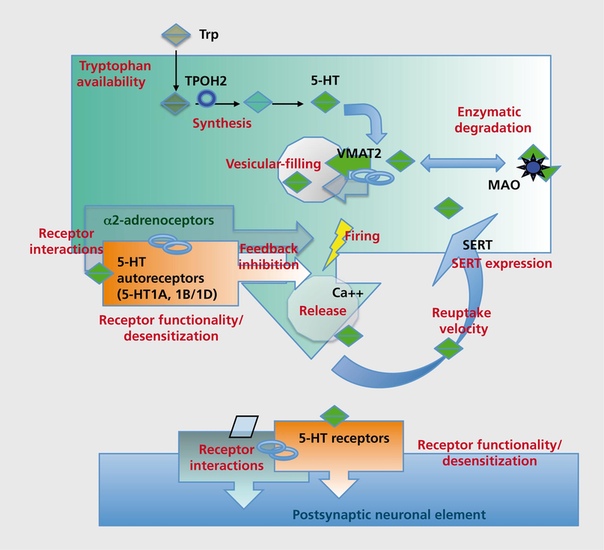
«Круговорот серотонина» легче всего представить по этой схеме:
Рецепторы серотонина
Существует 7 типов серотониновых рецепторов. Все они, за исключением 3-го типа, ионотропного, являющегося лиганд-зависимым ионным каналом, суть метаботропные. Также стоит отметить, что эти рецепторы при активации могут выполнять как тормозящую, так и возбуждающую функцию. При активации серотониновых рецепторов нейрон может выбрасывать не только различные низкомолекулярные нейромедиаторы (собственно серотонин, ацетилхолин, ГАМК, глутамат, дофамин, адреналин, норадреналин, октопамин), но и сигнальные пептиды и нейростероиды (кортизол, окситоцин, вазопрессин, пролактин, АКТГ, вещество Р и нейрокинины). Серотонинэргическая сигнальная система участвует в регуляции некоторых жизненно важных функций организма: сон, терморегуляция, пищевое поведение, когнитивно-мнестические способности, работа ЖКТ, тонус сосудов. Соответственно количеству функций эта система является мишенью множества фармакологически активных веществ: психотомиметиков/антипсихотиков, антидепрессантов, противорвотных, прокинетиков.
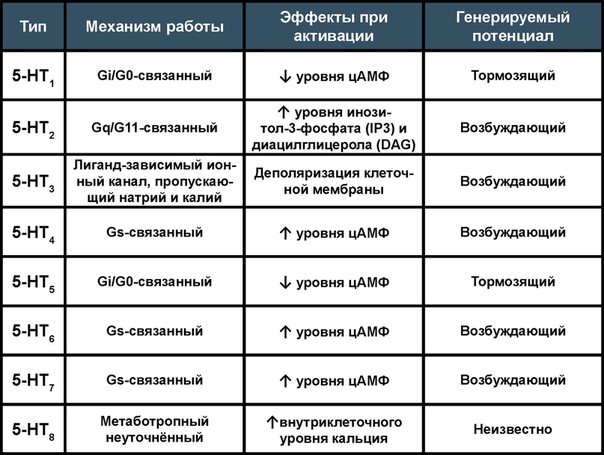
Стоит отметить, что 5-НТ8 рецептор у людей не встречается и был найден лишь у нескольких насекомых: бабочки репницы (Pieris rapae), хрущака малого булавоусого (Tribolium castaneum) и малярийных комаров вида Anopheles gambiae.
Серотониновые рецепторы типа 5-НТ1
Подтип 5-НТ1A

Иногда эти рецепторы называются «тормозящими».
При их активации происходит снижение артериального давления, снижается частота сердечных сокращений. Также эти рецепторы участвуют в механизмах регуляции тонуса .
Интересно, что влияние активации 5-НТ1А рецептора на последующий выброс нейромедиаторов различно у разных видов – вот пример, который стоит учитывать при разработке и тестировании лекарственных веществ: у грызунов этот подтип серотониновых рецепторов при активации увеличивает выброс норадреналина, в то время как у приматов – уменьшает. Однако в то же время у приматов (и людей в частности), активация 5-НТ1А увеличивает выброс дофамина в определённых областях мозга: полосатом теле, медиальной области префронтальной коры и гиппокампе.
Нейроны, содержащие на себе 5-НТ1А-рецепторы, расположены в самых разных отделах нервной системы: кора больших полушарий, гиппокамп, прозрачная перегородка, миндалевидное тело – в этих участках наблюдается наибольшая плотность вышеупомянутых рецепторов, однако их можно найти и в других местах: в базальных ганглиях и таламусе.
Многие агонисты 5-НТ1А рецепторов применяются в клинической практике в качестве анксиолитиков и антипсихотиков. Кроме того, в меньших дозировках, они назначаются в качестве корректоров при приёме антидепрессантов. На счёт антидепрессантов – существуют теории, что, помимо изменения экспрессии BDNF, за отсроченное наступление терапевтических эффектов ответственен механизм изменения плотности расположения 5-НТ1А рецепторов: за 2-3 недели приёма антидепрессантов уменьшается плотность пресинаптических ауторецепторов и повышается плотность постсинаптических рецепторов этого типа. Таким образом достигается двойной эффект: снижение тормозящего действия на пресинаптический нейрон, который начинает выбрасывать больше серотонина, и повышение чувствительности постсинаптического нейрона к собственно серотонину. Следует также иметь ввиду, что большая часть 5-НТ1А рецепторов – пресинаптические ауторецепторы, т.е. в большинстве своём не являются прямой целью релизеров серотонина, которые осуществляют своё действие через иные механизмы. И как уже указывалось выше, именно из-за этого существует такой разрыв во времени начала действия антидепрессантов разных классов – для развития действия селективных ингибиторов обратного захвата серотонина (СИОЗС) требуется время, а релизеры начинают своё действие очень быстро, однако оно длится не долго.
Кроме того, 5-НТ1А участвуют в регуляции ноцицепции (восприимчивости к боли): в нейронах, расположенных в дорсальной части ядер шва, эти рецепторы экспрессируются на одних и тех же клетках вместе с рецепторами тахикининов (NK1), при активации тормозя действие и последующий выброс вещества Р. Таким образом, области действия некоторых 5-НТ1А-агонистов и NK1-блокаторов могут перекрываться – оба класса веществ могут оказывать анальгезирующее и противорвотное действие.
Однако, не всё так хорошо: из-за замедления процессов высвобождения глутамата и ацетилхолина в гиппокампе, 5-НТ1А-агонисты при длительном приёме способны вызвать нарушения процессов запоминания. Некоторые блокаторы 5-НТ1А исследуются в качестве лекарств для улучшения памяти у пациентов с болезнью Альцгеймера. Интересно, что в других областях мозга, а именно префронтальной коре, этот тип рецепторов при активации, наоборот, увеличивает высвобождение ацетилхолина и дофамина, таким образом увеличивая способность к обработке информации, но не запоминанию.
При селективной активации 5-НТ1А-рецепторов наблюдаются следующие эффекты: снижение агрессии и импульсивности, повышение общительности и уровня эмпатии, снижение влечения к наркотикам или иным веществам, вызывающих позитивный дофаминовый отклик, ухудшение эрекции, снижение аппетита, продление фазы быстрого сна. Также эксперименты на животных выявили способность селективных 5-НТ1А-агонистов восстанавливать самостоятельное дыхание в случае передозировки опиоидами.
Этот тип серотониновых рецепторов также играет роль в регуляции выброса таких гормонов, как кортизол, кортикостерон, АКТГ, окситоцин, пролактин, соматотропный гормон, бета-эндорфин. Считается, что выброс окситоцина ответственен за анксиолитические эффекты при селективной активации, а выброс бета-эндорфина – за облегчение болевых ощущений.
Лиганды 5-НТ1А. Существует больше сотни различных веществ (агонистов/антагонистов), обладающих различной силой связывания и селективностью. Однако, в экспериментальной нейробиологии в качестве стандартного агониста используется 8-OH-DPAT.
Мутации. В настоящее время известно несколько десятков мутаций гена HTR1A, кодирующего этот рецептор, в основном это SNP, ведущие к замене единичной аминокислоты [7].
Подтип 5-НТ1B
Серотониновые рецепторы этого подтипа расположены главным образом в лобной доле коры больших полушарий, базальных ганглиях, полосатом теле и гиппокампе. Функции 5-НТ1В-рецепторов различаются в зависимости от расположения, например, во фронтальной области коры больших полушарий они являются постсинаптическими рецепторами, при активации тормозящими дальнейший выброс дофамина. Однако в базальных ганглиях и полосатом теле 5-НТ1В-рецептор является пресинаптическим ауторецептором, блокирующим выброс серотонина и глутамата. Интересно, что уровни экспрессии генов, кодирующих 5-НТ1В и 5-НТ1Е-рецепторы, взаимосвязаны, и, возможно, регулируются одним и тем же механизмом [8].
Вне ЦНС при активации 5-НТ1В-рецептор вызывает сужение сосудов, в основном в лёгких, однако он является также и мишенью суматриптана – противомигренозного средства. Кроме того, этот рецептор обнаруживается в молодых остеобластах и, согласно некоторым теориям, его активация в этих клетках служит сигналом для созревания, а в масштабах целого организма – к набору костной массы.
В опытах на мышах выявлено, что особи без нормально функционирующего 5-НТ1В-рецептора больше склонны к агрессии. Кроме того, мутантный фенотип практически невосприимчив к действию СИОЗС.
Подтип 5-НТ1C – данный подтип переименован в 5-НТ2C после детального изучения структуры и функций и, соответственно, отнесён в иную группу серотониновых рецепторов, которая будет рассмотрена ниже.
Подтип 5-НТ1D
Данный подтип отвечает за координацию движений, реакции стресса, пищевого поведения[9] и участвует в регуляции тонуса сосудов мозга. Высокая плотность рецепторов 5-НТ1D обнаруживается по ходу тройничного нерва.
Благодаря тому, что этот рецептор является одной из мишеней суматриптана и некоторых иных противомигренозных препаратов, его глубокое изучение помогло лучше понять механизмы нейрогенного воспаления. В дальнейшем выяснилось, что суматриптан, являясь лигандом тормозящего 5-НТ1D-рецептора, подавляет выброс CGRP (кальцитонин-ген родственный пептид), вещества Р и нейрокинина А, которые ответственны за возникновение болевых ощущений.
Подтип 5-НТ1Е
Изучение функций и маппинг (установление расположения) данного подтипа серотониновых рецепторов крайне затруднено в связи с отсутствием высокоселективных лигандов и антител. Единственный доступный лиганд, BRL-54443, не проявил достаточного уровня специфичности, т.к. является смешанным 5-HT1E/F-агонистом. Однако, методом исключения выявлена повышенная экспрессия 5-НТ1Е в лобной доле коры и гиппокампе, что позволяет предположить его участие в процессах запоминания[10].
Полный набор функций, регулируемых в организме данным рецептором, в настоящий момент возможно изучать лишь генетическим реверс-инжинирингом, сопоставляя мутации гена, кодирующего его, с наблюдаемыми отклонениями [11].
Подтип 5-НТ1F
Из-за высокой гомологии (более 57%) с предыдущим подтипом его изучение также было затруднено по причине почти полного отсутствия селективных лигандов. Однако, в сноске из описания предыдущего подтипа есть ссылка на статью с описанием синтеза и методики скрининга новых соединений, обладающих высокой селективностью и к 5-HT1F-рецептору, одним из которых оказался 5-N-бутирилокси-ДМТ. В настоящее время высокая плотность 5-НТ1F-рецепторов обнаружена в таких областях мозга, как скорлупа, гиппокамп и хвостатое ядро.
Серотониновые рецепторы типа 5-НТ2
В отличие от первого типа, они являются возбуждающими. Кроме того, эти рецепторы являются мишенью большинства психоактивных соединений, в т.ч. и использующихся в медицине.
Подтип 5-НТ2А
Как я уже упоминал выше, изначально серотониновые рецепторы делились на Д- и чувствительные, однако в англоязычной литературе под (Д) понимается дибензилин (он же феноксибензамин, альфа-1 адреноблокатор), в то время как в отечественной литературе (даже в такой авторитетной книге как «Фармакологическая регуляция психических процессов» за авторством Лаврецкой) под (Д) понимается ДЛК, он же ЛСД. Однако, как ни странно, суть от этого не меняется. Впоследствии, с развитием молекулярной биологии, когда стало возможно более подробно классифицировать рецепторы серотонина, Д-чувствительные были переименованы в 5-НТ2А.
Этот подтип серотониновых рецепторов обнаруживается в следующих областях ЦНС: неокортекс (префронтальная и теменная доли, также обнаруживается в постцентральной извилине, отвечающей за телесные ощущения), обонятельный бугорок, апикальные дендриты пирамидальных клеток 5 слоя коры больших полушарий. Выяснено, что 5-НТ2А-рецепторы способны взаимодействовать (оказывать модулирующее действие) со следующими типами рецепторов: 5-НТ1А, ГАМК-А, А1-рецепторы аденозина, АМПА, mGluR2, mGluR3, mGluR5, OX2 (рецепторы орексина). Взаимодействие с большим количеством глутаматных рецепторов (АМПА, mGluR2, mGluR3, mGluR5) не случайно, т.к. 5-НТ2А усиливает выброс глутамата за счёт своего возбуждающего действия[12].

Источник: http://www.frontiersin.org/
Помимо ЦНС 5-НТ2А-рецепторы обнаруживаются в фибробластах, тромбоцитах, клетках сердца и различных отделах ПНС.
Механизм работы 5-НТ2А-рецептора. При активации рецептора бета- и гамма-субъединицы «высвобождают» Gq-субъединицу, активирующую фосфолипазу С (PLC), которая, в свою очередь, превращает фосфатидилинозитолбисфосфат (PIP2) в диацилглицерол (DAG) и инозитолтрифосфат (IP3). DAG активирует протеинкиназу С (PKC), а IP3 запускает кальмодулин-зависимый механизм высвобождения кальция из эндоплазматического ретикулума. Существуют также побочные биохимические пути связанные с образованием арахидоновой кислоты из DAG. «Выбор» пути, по которому будут работать вторичные мессенджеры, может также зависеть от лиганда: например, если рассматривать DOB и 2C-B, то DOB (за счёт наличия метильной группы в альфа-положении на боковой цепи) действует по «основному» пути, описанному выше, а 2C-B – по «альтернативному», ведущему к повышению концентрации арахидоновой кислоты. Подробнее этот вопрос рассмотрен в лекции Дэвида Николса «Advances In Understanding How Psychedelics Work In The Brain».

Функции
В ЦНС: возбуждение, агрессия, участие в процессах запоминания[14]. Мышцы: сокращение гладкой мускулатуры кишечника, бронхов и сосудов. Участие в процессе свёртывания крови (прямое серотонилирование будет рассмотрено ниже).
Интересно, что относительно недавно было открыто участие серотонина в процессах воспаления, причём не только нейрогенного – известный галлюциноген DOI в дозировках, не влияющих на психический статус, показал способность подавлять ФНО-индуцированное воспаление[15]. Интересно, что такую же способность открыли и у ЛСД[16].
При активации 5-НТ2А-рецепторов в гипоталамусе в крови повышается уровень окситоцина, пролактина, АКТГ, кортикостерона и ренина.
Что касается лигандов данного вида рецепторов, то тут дело обстоит достаточно интересно: поскольку он является одной из главных мишеней огромного количества психоактивных веществ, начиная от ЛСД и мескалина, заканчивая экзотическими соединениями, которые лишь отдалённо похожи на ФЭА/триптамины/эрголины, то вполне логично, что он стал объектом внимания химиков-фармакологов, как официальных, так и не очень. В настоящее время известные базы, например isomerdesign, содержат сведения о нескольких сотнях веществ, тем или иным образом взаимодействующих с 5-НТ2А-рецепторами.
Если говорить о «классических» психоактивных веществах (псилоцин, псилоцибин, МДМА), то они в первую очередь они воздействуют на пирамидальные клетки префронтальной зоны коры – для возникновения повышенного уровня возбуждения нейронов данной области, которое обычно выражается в психозе с галлюцинациями, необходимо наличие рецепторного комплекса – димера из 5-НТ2А и mGluR2-рецепторов[18], который часто встречается именно в этой области мозга.
В гене, кодирующем данный рецептор, найдено (на текущий момент) более двух сотен различных мутаций[19]. К большому сожалению, большая часть публикаций по теме содержит результаты, которые лишь немного превышают статическую погрешность. Более того, результаты в некоторых публикациях довольно противоречивы[20]. Однако его роль в развитии психических отклонений трудно переоценить.
Подтип 5-НТ2В
Здесь ситуация выглядит несколько менее запутанной. Скорее всего, это связано с тем, что данный вид рецепторов не является мишенью весёлых и интересных химических веществ, являющихся причиной вдохновения у исследователей.
5-НТ2В-рецепторы обнаруживаются в следующих тканях и органах[21]: гладкие мышцы двенадцатипёрстной кишки, пищевода, тонкого и толстого кишечника; сердце, лёгкие (гладкие мышцы бронх и лёгочных артерий), кора надпочечников, корковое вещество почек, мозжечок (клетки Пуркинье, клетки зернистого слоя коры мозжечка, вставочные клетки), конечный мозг (гиппокамп).
Помимо своего влияния на поведение (он ответственен за двигательное возбуждение при введении психоактивных веществ), он также влияет на сердечно-сосудистую систему. При возбуждении 5-НТ2В-рецепторов, расположенных вне мозга, наблюдается сужение сосудов и повышение АД. Согласно данным исследований, это действие прямое, неопосредованное, т.к. введение бета-адреномиметиков крысам, у которых 5-НТ2В-рецептор уже заблокирован, не приводило к повышению АД. Кроме того, было выявлено, что 5-НТ2В-агонисты способны вызывать фиброз сердца при длительном приёме – это, опять же, связано с прямым участием 5-НТ2В-рецепторов в запуске процесса пролиферации фибробластов. С этим механизмом может быть связана часть смертельно опасных побочных эффектов психоделика Bromo-dragonfly.
Кстати, помимо прямого пролиферативного действия на фибробласты, активация этих рецепторов может также помочь в регенерации клеток печени[22].
Интересно, что при «серотониновом синдроме» (опасном побочном эффекте некоторых антидепрессантов, выражающемся в бесконтрольном высвобождении серотонина и опасном повышении его концентрации во внеклеточной среде) данный подтип рецепторов оказывает защитное действие[23].
Подтип 5-НТ2C
Этот вид серотониновых рецепторов сначала был отнесён к группе 5-НТ1, потом его признали тем же рецептором, что и 5-НТ2А (из-за высокой гомологии), и только относительно недавно он был выделен в отдельный подтип.
5-НТ2С-рецептор обнаруживается (ИГХ + анализ мРНК в тканях[24]) в сосудистом сплетении желудочков мозга, в пятом слое коры больших полушарий (за исключением шпоровидной борозды), мозолистом теле, септальных ядрах, прилежащем ядре, базальных ядрах переднего мозга, хвостатом ядре, скорлупе, диагональной полоске (полоске Брока), вентральном стриатуме, частично – миндалевидном теле, среднем мозге, таламусе, ядрах ствола мозга. В основном на нейронах обнаруживается соматодендритная локализация 5-НТ2С-рецептора.
Данный рецептор при активации замедляет выброс дофамина и норадреналина, однако в некоторых областях мозга он также регулирует выброс ацетилхолина. Интересно, что в связи с этим, 5-НТ2С-блокаторы исследуются в качестве потенциальных новых антидепрессантов. Также, если рассматривать старые классические СИОЗС, например флуоксетин, то они, наоборот, в первые 2 недели приёма косвенно повышают тормозящее действие 5-НТ2С-рецепторов, которое потом сглаживается их down-регуляцией, т.е. «понижением чувствительности» — понижением плотности расположения рецепторов на клеточной мембране в ответ на продолжительную их стимуляцию. За счёт тормозящего действия 5-НТ2С-рецептора на выброс дофамина, его блокаторы и обратные агонисты косвенно проявляют дофаминомиметическое действие.
Интересно, что повышение уровня цитокинов влечёт за собой повышение уровня экспрессии гена 5-НТ2С-рецептора. Этот факт может служить одним из объяснений изменения психического статуса при различных хронических воспалениях.
5-НТ2С-рецепторы принимают участие в регуляции работы органов гипоталамо-гипофизарно-надпочечниковой системы. Обнаружено прямое повышение уровня КРГ и вазопрессина при стимуляции серотониновых рецепторов паравентрикулярных ядер гипоталамуса.
Так как ген HTR2C расположен на Х-хромосоме, то мужчины несут одну копию гена, а женщины – две. Это также влияет на работу серотонинэргической системы в целом, т.к. в данном случае проявляется «мозаицизм» — в каждой клетке будет работать только одна Х-хромосома несущая один из двух вариантов гена. Кроме того, вне зависимости от пола, мРНК этого рецептора подвергается довольно редкому для генов этого семейства виду модификации – выборочной замены аденозина на инозин с помощью мозгоспецифичного фермента ADAR3. В процессе трансляции инозин равнозначен гуанозину. Существует несколько сайтов редактирования мРНК, чьё случайное изменение может дать 24 различных изоформы рецептора. Это отражается на функциях рецептора – так называемая VGV-изоформа, где из-за редактирования мРНК в конечной аминокислотной последовательности белка на 157 и 161 позициях стоит валин, а на 159 – глицин, значительно менее активна и вызывает меньший отклик в повышении концентрации вторичных мессенджеров при одинаковых концентрациях серотонина и различных агонистов, нежели не редактированная INI-изоформа (по 157 и 161 позициям – изолейцин, по 159 – аспарагиновая кислота). Интересно, что в некоторых исследованиях в области психиатрии и генетики обнаруживается связь с расстройствами не только у лиц, у которых выявлена мутация гена рецептора, но также и у тех, у кого нарушены процессы редактирования мРНК в мозге[25].
Серотониновые рецепторы типа 5-НТ3
Данный вид рецепторов является уникальным в своём роде – это единственный не-метаботропный тип. 5-НТ3-рецептор представляет собой лиганд-зависимый ионный канал, состоящий из пяти различных субъединиц (5-HT3A, 5-HT3В, 5-HT3С, 5-HT3D, 5-HT3E). Он может быть представлен в виде гомопентамера, состоящего исключительно из одних 5-НТ3А-субъединиц, либо гетероментамера – когда в его составе, помимо обязательно наличествующей 5-НТ3А-субъединицы, находятся субъединицы других подтипов. Интересно, что по уровню гомологии субъединицы можно разделить на 2 группы: первая включает в себя 5-HT3A и 5-HT3В, вторая 5-HT3С, 5-HT3D и 5-HT3E.
Каждая субъединица состоит из четырёх участков(М1-М4), представляющих собой аминокислотную цепь, уложенную в форме альфа-спиралей, которые пересекают клеточную мембрану. N-конец аминокислотной цепи субъединицы локализован на внешней стороне клеточной мембраны и осуществляет связывание лигандов, трансмембранные участки (а конкретнее – аминокислотная петля, связывающая М2 и М3-участки) непосредственно задействованы в механизме открытия ионного канала. Также присутствует внутриклеточная петля между М3 и М4-участками, участвующая в транспортировке и регуляции работы субъединицы.
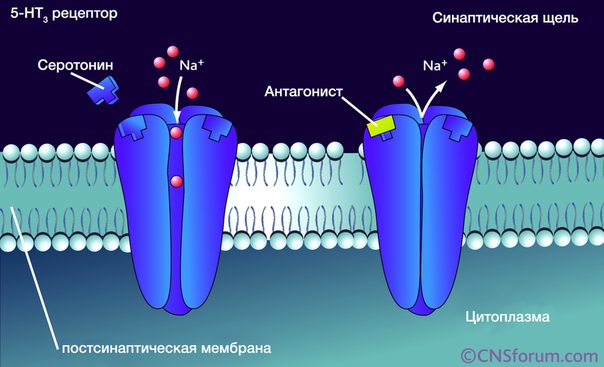
Рецептор обнаруживается в тонком и толстом кишечнике, миндалевидном теле, гиппокампе, хвостатом ядре, вставочных нейронах неокортекса, тимусе, селезёнке, предстательной железе.
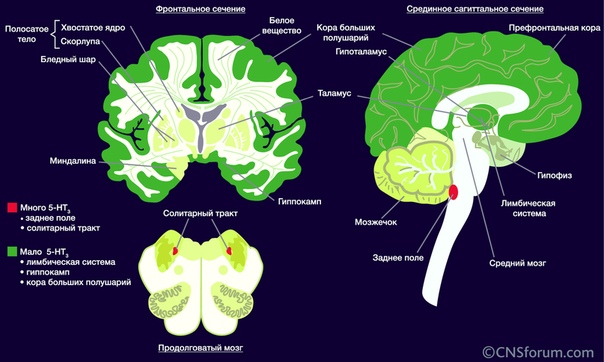
Помимо регуляции высшей нервной деятельности, данный тип ионотропных рецепторов серотонина играет важную роль в запуске рвотного рефлекса. Это было выявлено почти случайно – врачи ещё в ХХ веке обнаружили противорвотные свойства некоторых антипсихотиков, и уже позже, с развитием молекулярной биологии, стала ясна причина такого действия – прямое подавление активности соответствующих ядер продолговатого мозга.
Вполне логично, что свойства рецептора обусловлены свойствами субъединиц, в него входящими:
5-НТ3А-субъединица. Гомопентамерные 5-НТ3-рецепторы, содержащие лишь 5-НТ3А-субъединицу, обладают меньшей проводимостью (0,4 pS (пикоСименс)), нежели гетеропентамерные, в которых одна из субъединиц заменена на 5-НТ3В (проводимость — 16 pS). Во время изучения вариантов мутации гена 5-НТ3А-субъединицы был найден участок цепи, названный «HA-stretch», при обогащении которого остатками аргинина, по типу 5-НТ3В-рецептора, общая проводимость рецептора возрастала в 28 раз.
5-HT3B-субъединица. Данная субъединица может формировать ионный канал только вместе с 5-НТ3А-субъединицами. Кроме того, исследования выявили роль содержащих её рецепторов в развитии депрессии: найден важный полиморфизм, заключающийся в замене тирозина в 129-й позиции аминокислотной цепи на серин, выражающейся в более медленной десенсибилизации рецептора и более длинным временем работы ионного канала при активации. Носители данной мутации менее склонны к депрессивным расстройствам[26].
5-НТ3С-субъединица. Рецепторы, её содержащие, обнаруживаются в мозге взрослого человека, прямой кишке, мышцах, лёгких, тонком кишечнике, на стадии внутриутробного развития они также присутствуют в почках и формирующемся кишечнике. Её мутации связывают с избыточным рвотным рефлексом, особенно при химиотерапии. Также были выявлены SNP, повышающие риск развития расстройств аутистического спектра.
5-НТ3D-субъединица. Присутствует в печени взрослого человека, в почках и прямой кишке взрослого и зародыша. Её мутации могут утяжелять течение шизофрении и также понижают порог рвотного рефлекса.
5-НТ3Е-субъединица. Изначально она была обнаружена лишь в тканях толстой кишки взрослого человека, однако потом незначительные количества содержащих ее 5-НТ3-рецепторов обнаружили в спинном мозге и сетчатке. Вполне возможно её наличие в мозге, т.к. вещества, избирательно блокирующие раскрытие 5-НТ3-рецепторов, содержащих 5-НТ3Е-субъединицы, показали способность облегчать течение обсессивно-компульсивного расстройства.
Серотониновые рецепторы типа 5-НТ4
Данный вид рецепторов является метаботропным, при активации повышающим количество цАМФ в клетке, что в свою очередь запускает каскад реакций, ведущий к выбросу нейромедиаторов. Ранее он считался «кардиоспецифичным[27]» рецептором, так как изначально был обнаружен в наибольших концентрациях в предсердных отделах проводящей системы сердца. Однако в дальнейшем 5-НТ4-рецепторы обнаружили и в других тканях и органах: ЖКТ, мочевой пузырь, надпочечники. В ЦНС: скорлупа, хвостатое ядро, прилежащее ядро, бледный шар, чёрное вещество, реже встречается в неокортексе, ядрах шва, Варолиевом мосту. Полностью отсутствуют в мозжечке. В стволе мозга 5-НТ4-рецепторы контролируют деятельность дыхательного центра. На мышах даже был проведён интересный эксперимент: им вводился фентанил в дозах, способных вызвать подавление самостоятельного дыхания, а затем вводились агонисты 5-НТ4-рецепторов (конкретнее – BIMU-8). В результате у подопытных животных восстановилось самостоятельное дыхание, но обезболевающее действие фентанила осталось[28]. Теоретически, данный эффект можно использовать для расширения «терапевтического окна» опиоидных анальгетиков.
Вклад мутаций гена этого рецептора в развитие шизофрении достаточно противоречив, однако их связь с возникновением биполярного расстройства подтверждёна достоверно.
Серотониновые рецепторы типа 5-НТ5
Среди всех рецепторов серотонина этот тип является наименее изученным. Пока известно лишь то, что в геноме человека представлено 2 подтипа – 5-НТ5А и 5-НТ5В. Однако, ген второго подтипа является псевдогеном, т.к. в его последовательности содержится стоп-кодон, что делает его нефункциональным. У грызунов гены обоих подтипов рецептора являются функциональными.
Касательно расположения – методом RT-PCR (ПЦР в реальном времени) установлено, что экспрессия функционального гена 5-НТ5А-рецептора наблюдается лишь в мозге. Кстати, исследователей, публикующих статьи на тему данного типа рецепторов, можно поблагодарить за то, что они публикуют и негативные результаты. Например, мутации 5-НТ5-рецептора не влияют на развитие тревожных расстройств. Однако, в некоторых публикациях содержатся указания на его возможную роль в развитии шизофрении (полиморфизм Pro-15-Ser, присутствие которого увеличивает риск возникновения данного заболевания) и регуляции циркадных ритмов. На сегодняшний день достоверно установлена связь мутаций этого рецептора с т.н. «исследовательским поведением» (exploratory behaviour), а проще говоря – любознательностью. Также исследования выявили, что у мышей, нокаутных по 5-НТ5А-рецептору, снижено двигательное возбуждение при введении ЛСД.
Кстати, несмотря на малую изученность, мы все хотя бы раз в жизни кушали блокаторы 5-НТ5-рецепторов. Я говорю о валерьяне лекарственной, которая содержит валереновую кислоту. Помимо позитивной аллостерической модуляции работы ГАМК-А-рецептора, это вещество также возбуждает серотониновые 5-НТ5-рецепторы, которые оказывают тормозящее действие на выброс нейромедиаторов, что также осуществляет вклад в анксиолитические и седативные эффекты экстракта валерианы.
Серотониновые рецепторы типа 5-НТ6
Рецепторы данного типа обнаружены исключительно в мозге: обонятельный бугорок, прилежащее ядро, кора больших полушарий, полосатое тело, хвостатое ядро, гиппокамп, зернистый слой мозжечка. Предполагается, что он участвует в регуляции процессов запоминания, координации движений и возникновении эмоций.
Несмотря на то, что рецепторы данного типа считаются возбуждающими (при активации увеличивают уровень цАМФ в цитоплазме нейрона), их влияние на выброс других нейромедиаторов неоднозначно: нейроны, содержащие 5-НТ6-рецепторы и расположенные во фронтальной области коры, при возбуждении специфическими агонистами выбрасывают большие количества ацетилхолина и глутамата, в то время как введение антагонистов увеличивает концентрацию дофамина и норадреналина. Интересно, что активация 5-НТ6-рецепторов, расположенных на ГАМКэргических нейронах, оказывает в целом тормозящий эффект, т.е. наблюдается выброс ГАМК.
Неоднозначное влияние данного рецептора на процессы в ЦНС упомянуто не зря, так как его агонисты, например EDMT, несмотря на стимуляцию выброса ацетилхолина и глутамата, ухудшали процессы запоминания у экспериментальных животных: подавлялась как кратковременная, так и долговременная память. Интересно, что антагонисты 5-НТ6-рецепторов исследуются в качестве перспективных лекарств от болезни Альцгеймера и других форм когнитивной недостаточности (в настоящее время выявлено несколько мутаций гена этого рецептора, значительно увеличивающих риск развития нейродегенеративных заболеваний), а агонисты – в качестве антидепрессантов и анксиолитиков. Новый 5-НТ6 агонист E-6837 в данный момент проходит испытания на животных в качестве средства для подавления аппетита, где он показал себя более эффективным и безопасным средством, нежели использующийся в настоящее время сибутрамин.
Серотониновые рецепторы типа 5-НТ7
Эти метаботропные серотониновые рецепторы также запускают процессы высвобождения нейромедиаторов путём запуска каскада реакций фосфорилирования через цепь вторичных мессенджеров, т.е. являются «возбуждающими». Изначально они были идентифицированы в качестве подтипа 5-НТ1-рецепторов, однако позже, благодаря изучению их структуры и аминокислотной последовательности, были выделены в отдельный тип.
Эти рецепторы регулируют тонус гладкой мускулатуры ЖКТ и сосудов, циркадные ритмы, процессы запоминания и обработки информации, также они участвуют в терморегуляции. Обнаруживаются в таламусе, гипоталамусе, неокортексе и гиппокампе. Интересно, что согласно последним исследованиям функции 5-НТ7-рецептора этим не ограничиваются. Более того, опыты показали, что, например, его активация может как усиливать, так и подавлять ноцицепцию. Не менее неоднозначным является факт, что мыши, нокаутные по гену этого рецептора, проявляют в опытах иное поведение, нежели животные с рецептором, заблокированным специфическими антагонистами! Бельгийские учёные в своей статье[29] предлагают следующее объяснение такому сильному различию в результатах экспериментов: при определённых условиях часть рецепторов является Gs-связанными, как и было установлено изначально при открытии 5-НТ7, однако ещё часть связывается с другими G-белками, реализуя своё действие через иные сигнальные каскады.
5-НТ7 может присутствовать в трёх различных вариантах, являющихся продуктом одного гена HTR7 и получающихся в результате альтернативного сплайсинга – 5-НТ7(а), 5-HT7(b) и 5-HT7(d). Между этими изоформами нет существенной разницы в связывании химических веществ – агонисты, как и антагонисты, имеют к ним одинаковый аффинитет. Различие заключается в уровнях, на которые повышается цАМФ при активации рецептора специфическими агонистами – изоформа 5-HT7(d) является наиболее активной[30].
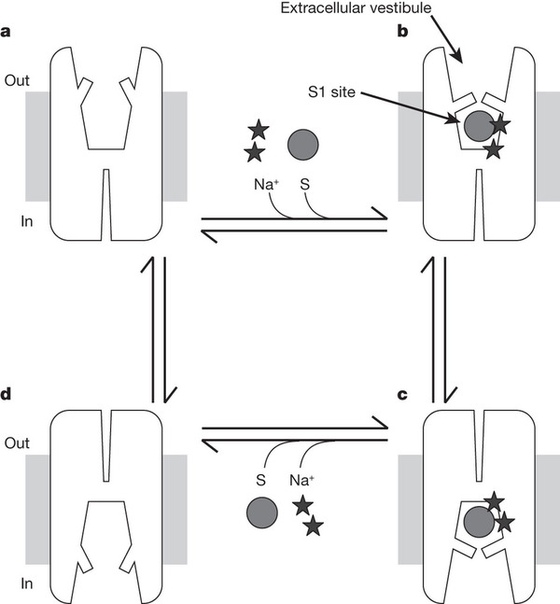
Серотониновый транспортёр (SERT)
Помимо рецепторов, не менее важным звеном серотонинэргической сигнализации является серотониновый транспортёр (SERT). Он осуществляет обратный захват нейромедиаторов из синапса обратно в клетку. Интересно, что многие представляют себе этот белок в качестве свободно находящегося в цитоплазме фермента (вроде моноаминоксидазы), который «цепляет» молекулы серотонина и закачивает их обратно. Такое представление в корне неверно:этот белок заякорен на клеточной мембране. Этот белок является Na/Cl-зависимым транспортёром, переносящим серотонин вместе с ионами натрия внутрь клетки.
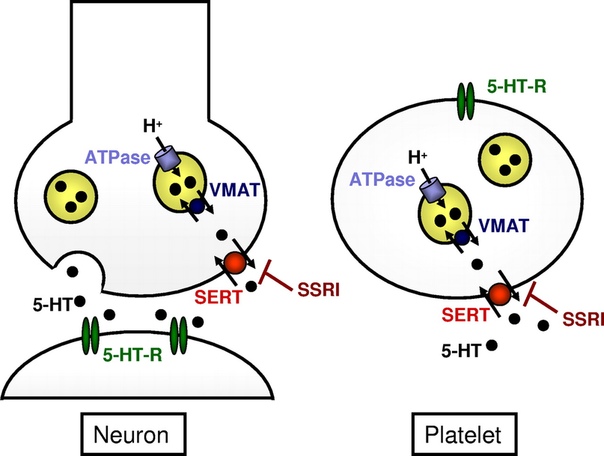
Он контролирует концентрацию серотонина, доступного для связывания с рецепторами. Неудивительно, что он является мишенью множества психоактивных веществ, например антидепрессантов. Подробнее механизм влияния концентрации серотонина на плотность рецепторов был уже рассмотрен выше. Стоит также отметить, что патологии (мутации, в т.ч. и SNP) связанные с этим белком-транспортером, влияют не только на вероятность развития различных психических заболеваний (шизофрения, ОКР, биполярное расстройство, депрессия), но и на выраженность ответа на их лечение[31]. Кстати, если читатель – белый европеоид, то ему повезло – он легче перенесёт лечение антидепрессантами группы СИОЗС (сертралин, пароксетин, флувоксамин, флуоксетин, эсциталопрам). Меньше повезло азиатам – у них переносимость СИОЗС гораздо ниже, т.е. имеется больший риск развития побочных эффектов, также снижена эффективность при лечении препаратами данной группы.
Роль серотонина в свертывании крови
Как уже было описано выше, специфические рецепторы присутствуют и на тромбоцитах. При их активации серотонином запускается каскад реакций вторичных мессенджеров. Ключевую роль в них играет G-белок, который является ключевым для дальнейших биохимических реакций, которые можно разделить несколько путей:
- Активация фактора обмена гуаниновых нуклеотидов p115 RhoGEF и лейкемия-ассоциированного фактора обмена нуклеотидов LARG. Они, в свою очередь, активируют ГТФазу RhoA. (Которая может быть активирована серотонином напрямую в ходе рецептор-независимого взаимодействия – серотонилирования, с помощью фермента трансглутаминазы. Интересно, что нахождение трансглутаминазы в тромбоцитах было установлено ещё 50 лет назад, в то время как её роль в переносе серотонина была выявлена относительно недавно). Она запускает работу Ro-ассоцированной киназы (ROCK), которая через промежуточные киназы LIMK1 и LIMK2 фосфорилирует кофилин (белок ответственный за деполимеризацию актина) и тем самым выключает его. Также ROCK путём фосфорилирования регуляторной субъединицы выключает работу MLCP (в некоторых источниках – MYPT1, фосфатаза лёгких цепей миозина), что выражается в снижении уровня фосфорилирования самого миозина. Все эти эффекты, направленные на регуляцию белков цитоскелета, происходят ради изменения формы тромбоцита, что необходимо для их агрегации в месте повреждения кровеносных сосудов.
- Параллельно протекает ещё один процесс. G-белок запускает работу фосфолипаз PLC-beta2/3. Фосфолипазы С расщепляют фосфатидилинозитолбисфосфат (PIP2) на диацилглицерол (DAG) и инозитолтрифосфат (IP3). В данный момент нам интересен лишь DAG (вместе с ионами кальция, см. ниже), который запускает протеинкиназу С (PKC-alpha). Эта протеинкиназа фосфорилирует синтаксин 4 (Syntaxin 4) и SNAP-23, которые, будучи в активном состоянии, вместе с белком VAMP3 запускают процесс высвобождения везикул с сигнальными веществами. Интересно, что эти везикулы содержат в том числе и сам серотонин, чьё повышение концентрации «рекрутирует» другие тромбоциты для участия в процессе образования тромба.
- Теперь обратимся к другом продукту работы фосфолипазы – IP3. Он, взаимодействуя с со специфическим рецептором инозитолтрифосфата (IP3R1), запускает высвобождение кальция из эндоплазматического ретикулума в цитозоль. Ионы кальция, помимо активации вышеупомянутой протеинкиназы С, участвуют в повышении сродства рецепторов белков клеточной адгезии (интегрины бета 3, альфа 2b; комплекс интегринов альфа 2b/бета 3) к фибриногену. Этот процесс необходим для эффективного «слипания» тромбоцитов между собой и образования тромба.
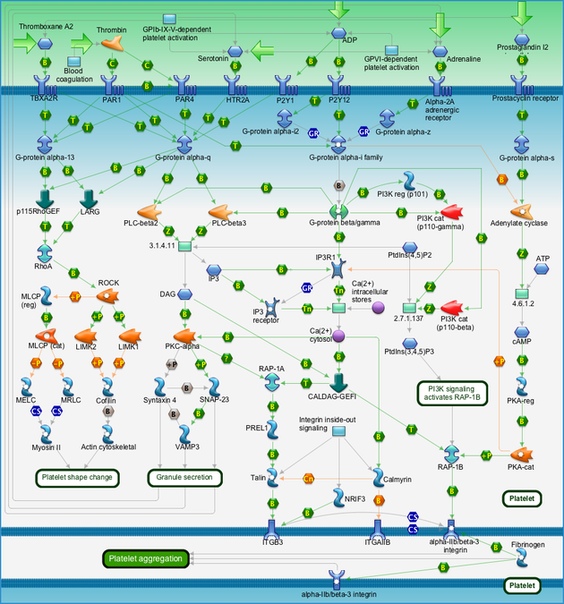
Источник: http://www.bio-rad.com/webroot/web/images/lsr/global/english/products/amplification_pcr/primepcr/pathway/zoom_out/2442_map.png
Заключение
Несомненно, я бы мог рассказать уважаемому читателю об увлекательных видениях и обманах восприятия, которые человеческий мозг генерирует будучи отравленым ЛСД или мескалином. Однако, согласно современным представлениям о природе галлюцинаций, их содержание не зависит от источника их вызвавшего. Конечно, в обсуждении этой статьи обязательно появятся люди, которые будут утверждать что под 25N-nBOMe к ним приходят красные эльфы, а под 25I-nBOMe – зелёные, но это будет лишь субъективным опытом, не относящимся к широкой выборке, и, скорее всего, зависящим от сеттинга трипа. В плацебо-контролируемых рандомизированных многоцентровых исследованиях было выявлено, что вышеупомянутый 25N-nBOMe у разных людей в разной обстановке даёт совершенно разные результаты: кто-то о либеральных реформах с портретом Парфёнова беседует, кто-то рыбок ловит, а кто-то ацтекские узоры на стенах ищет.
Кроме шуток. Многие психоактивные вещества, которые используются в медицине для лечения различных заболеваний, не являются продуктом каких-нибудь сложных SAR-расчётов, просто так звёзды сложились – внимательные клиницисты начали обращать внимание на состояние пациентов после введение противовоспалительных, сосудосуживающих или антигистаминных веществ. Сейчас, для того, чтобы разработать новое лекарство от депрессии или грозный хардкорный психоделик, валящий с ног ветеранов DOB-оедов, не нужно несколько лет стоять за колбами и пробирками – достаточно иметь данные о трёхмерной структуре рецептора и сайтах связывания, голову на плечах и немного воображения.
Я надеюсь, что эта статья выйдет чуточку ранее 19 апреля – знаменательного для для всей психофармакологии и нейрофизиологии, когда Альберт Хоффман испытал на себе эффекты ЛСД. Именно 19 апреля миру было показано, что 100 микрограмм вещества, которые даже невооружённым взглядом увидеть трудно, способны изменить ход мыслей одного человека и ход истории для миллионов.
[1] http://trv-science.ru/2009/03/03/shizofreniya-kak-posledstvie-ehvolyucii-mozga/
[2] P.Khaitovich et al. Metabolic changes in schizophrenia and human brain evolution. Genome Biology. 2008. 9: R124. http://genomebiology.com/2008/9/8/R124
[3] Glick SD, Maisonneuve IM, Szumlinski KK (2001). «Mechanisms of action of ibogaine: relevance to putative therapeutic effects and development of a safer iboga alkaloid congener». Alkaloids Chem Biol. The Alkaloids: Chemistry and Biology 56: 39–53.
[4] Walther DJ, Peter JU, Winter S, H?ltje M, Paulmann N, Grohmann M, Vowinckel J, Alamo-Bethencourt V, Wilhelm CS, Ahnert-Hilger G, Bader M. (2003). Serotonylation of small GTPases is a signal transduction pathway that triggers platelet alpha-granule release. Cell. 115(7):851-62
[5] Serotonin and its metabolism in basal deuterostomes: insights from Strongylocentrotus purpuratus and Xenoturbella bocki
Leah N. Squires, Stanislav S. Rubakhin, Andinet Amare Wadhams, Kristen N. Talbot, Hiroaki Nakano, Leonid L. Moroz, Jonathan V. Sweedler
Journal of Experimental Biology 2010 213: 2647-2654; doi: 10.1242/jeb.042374
[6] Qi YX, Xia RY, Wu YS, Stanley D, Huang J, Ye GY (2014). «Larvae of the small white butterfly, Pieris rapae, express a novel serotonin receptor». J. Neurochem. 131: 767–77. doi:10.1111/jnc.12940. PMID 25187179.
[7] Drago A, Ronchi DD, Serretti A (August 2008). «5-HT1A gene variants and psychiatric disorders: a review of current literature and selection of SNPs for future studies». Int. J. Neuropsychopharmacol. 11 (5): 701–21. doi:10.1017/S1461145707008218. PMID 18047755.
[8] Shoval G, Bar-Shira O, Zalsman G, John Mann J, Chechik G (Jul 2014). «Transitions in the transcriptome of the serotonergic and dopaminergic systems in the human brain during adolescence». European Neuropsychopharmacology 24 (7): 1123–32. doi:10.1016/j.euroneuro.2014.02.009. PMID 24721318.
[9] «Further evidence of association of OPRD1 & HTR1D polymorphisms with susceptibility to anorexia nervosa.» Brown KM, Bujac SR, Mann ET, Campbell DA, Stubbins MJ, Blundell JE. Biol Psychiatry. 2007 Feb 1;61(3):367-73. Epub 2006 Jun 27.PMID:16806108
[10] J Pharmacol Exp Ther. 2011 Jun;337(3):860-7. doi: 10.1124/jpet.111.179606. Epub 2011 Mar 21.
Toward selective drug development for the human 5-hydroxytryptamine 1E receptor: a comparison of 5-hydroxytryptamine 1E and 1F receptor structure-affinity relationships.
Klein MT1, Dukat M, Glennon RA, Teitler M.
[11] Am J Med Genet B Neuropsychiatr Genet. 2003 Apr 1;118B(1):36-40.
Suicide and serotonin: study of variation at seven serotonin receptor genes in suicide completers.
Turecki G1, Sequeira A, Gingras Y, S?guin M, Lesage A, Tousignant M, Chawky N, Vanier C, Lipp O, Benkelfat C, Rouleau GA.
[12] Zhang C, Marek GJ (January 2008). «AMPA receptor involvement in 5-hydroxytryptamine2A receptor-mediated pre-frontal cortical excitatory synaptic currents and DOI-induced head shakes». Progress in Neuro-psychopharmacology & Biological Psychiatry 32 (1): 62–71. doi:10.1016/j.pnpbp.2007.07.009. PMID 17728034.
[13] https://vimeo.com/16782003
[14] Harvey JA (2003). «Role of the serotonin 5-HT(2A) receptor in learning». Learn. Mem. 10 (5): 355–62. doi:10.1101/lm.60803. PMC 218001. PMID 14557608.
[15] Nau F, Yu B, Martin D, Nichols CD (2013). «Serotonin 5-HT2A Receptor Activation Blocks TNF-a Mediated Inflammation In Vivo». PLOS ONE 8 (10): e75426. doi:10.1371/journal.pone.0075426. PMID 24098382.
[16] Yu B, Becnel J, Zerfaoui M, Rohatgi R, Boulares AH, Nichols CD (November 2008). «Serotonin 5-hydroxytryptamine(2A) receptor activation suppresses tumor necrosis factor-_-induced inflammation with extraordinary potency». J. Pharmacol. Exp. Ther. 327 (2): 316–323. doi:10.1124/jpet.108.143461. PMID 18708586.
[17] Moreno JL, Muguruza C, Umali A, Mortillo S, Holloway T, Pilar-Cu?llar F, Mocci G, Seto J, Callado LF, Neve RL, Milligan G, Sealfon SC, L?pez-Gim?nez JF, Meana JJ, Benson DL, Gonz?lez-Maeso J (2012). «Identification of Three Residues Essential for 5-Hydroxytryptamine 2A-Metabotropic Glutamate 2 (5-HT2A{middle dot}mGlu2) Receptor Heteromerization and Its Psychoactive Behavioral Function». Journal of Biological Chemistry 287 (53): 44301–44319. doi:10.1074/jbc.M112.413161. PMC 3531745. PMID 23129762.
[18] Moreno JL, Holloway T, Albizu L, Sealfon SC, Gonz?lez-Maeso J (2011). «Metabotropic glutamate mGlu2 receptor is necessary for the pharmacological and behavioral effects induced by hallucinogenic 5-HT2A receptor agonists». Neuroscience Letters 493 (3): 76–79. doi:10.1016/j.neulet.2011.01.046. PMC 3064746. PMID 21276828.
[19] Chambers JJ, Kurrasch-Orbaugh DM, Parker MA, Nichols DE (March 2001). «Enantiospecific synthesis and pharmacological evaluation of a series of super-potent, conformationally restricted 5-HT(2A/2C) receptor agonists». Journal of Medicinal Chemistry 44 (6): 1003–10. doi:10.1021/jm000491y. PMID 11300881.
[20] Serretti A, Drago A, De Ronchi D (2007). «HTR2A gene variants and psychiatric disorders: a review of current literature and selection of SNPs for future studies». Current medicinal chemistry 14 (19): 2053–69. doi:10.2174/092986707781368450. PMID 17691947.
[21] FEBS Lett. 1996 Aug 5;391(1-2):45-51. «Immunohistochemical localisation of the serotonin 5-HT2B receptor in mouse gut, cardiovascular system, and brain» Choi DS1, Maroteaux L.
[22] Ebrahimkhani MR, Oakley F, Murphy LB, Mann J, Moles A, Perugorria MJ, Ellis E, Lakey AF, Burt AD, Douglass A, Wright MC, White SA, Jaffr? F, Maroteaux L, Mann DA (Dec 2011). «Stimulating healthy tissue regeneration by targeting the 5-HT?B receptor in chronic liver disease». Nature Medicine 17 (12): 1668–73. doi:10.1038/nm.2490. PMC 3428919. PMID 22120177.
[23] Diaz SL, Maroteaux L (Sep 2011). «Implication of 5-HT(2B) receptors in the serotonin syndrome». Neuropharmacology 61 (3): 495–502. doi:10.1016/j.neuropharm.2011.01.025. PMID 21277875.
[24] Synapse. 2001 Oct;42(1):12-26. «Regional distribution and cellular localization of 5-HT2C receptor mRNA in monkey brain: comparison with [3H]mesulergine binding sites and choline acetyltransferase mRNA.» L?pez-Gim?nez JF1, Mengod G, Palacios JM, Vilar? MT.
[25] Iwamoto K, Kato T (August 2003). «RNA editing of serotonin 2C receptor in human postmortem brains of major mental disorders». Neurosci. Lett. 346 (3): 169–72. doi:10.1016/S0304-3940(03)00608-6. PMID 12853111.
[26] Krzywkowski, K., Davies, P. A., Feinberg-Zadek, P. L., Brauner-Osborne, H., Jensen, A. A. High-frequency HTR3B variant associated with major depression dramatically augments the signaling of the human 5-HT(3AB) receptor. Proc. Nat. Acad. Sci. 105: 722-727, 2008. PubMed: 18184810
[27] Blondel, O., Vandecasteele, G., Gastineau, M., Leclerc, S., Dahmoune, Y., Langlois, M., Fischmeister, R. Molecular and functional characterization of a 5-HT(4) receptor cloned from human atrium. FEBS Lett. 412: 465-474, 1997. [PubMed: 9276448] [28] Manzke, T., Guenther, U., Ponimaskin, E. G., Haller, M., Dutschmann, M., Schwarzacher, S., Richter, D. W. 5-HT4(a) receptors avert opioid-induced breathing depression without loss of analgesia. Science 301: 226-229, 2003. [PubMed: 12855812] [29] Role of the 5-HT7 receptor in the central nervous system: from current status to future perspectives. Matthys A, et al. Mol Neurobiol, 2011 Jun. PMID 21424680
[30] Krobert KA and Levy FO (2002) The human 5-HT7 serotonin receptor splice variants: constitutive activity and inverse agonist effects. Br J Pharmacol 135(6):1563-1571. (2002). «The human 5-HT7 serotonin receptor splice variants: constitutive activity and inverse agonist effects.». British Journal of Pharmacology 135 (6): 1563–71. doi:10.1038/sj.bjp.0704588. PMC 1573253. PMID 11906971.
[31] Acta Clin Croat. 2013 Sep;52(3):353-62.
Serotonin transporter gene (5-HTTLPR) polymorphism and efficacy of selective serotonin reuptake inhibitors—do we have sufficient evidence for clinical practice.
Karlovi? D1, Karlovi? D2.
Телеграм: t.me/ainewsline
Источник: m.vk.com