Половое размножение: преимущества и недостатки
МЕНЮ
Главная страница
Поиск
Регистрация на сайте
Помощь проекту
Архив новостей
ТЕМЫ
Новости ИИ
Городские сумасшедшие
ИИ в медицине
ИИ проекты
Искусственные нейросети
Искусственный интеллект
Слежка за людьми
Угроза ИИ
ИИ теория
Компьютерные науки
Машинное обуч. (Ошибки)
Машинное обучение
Машинный перевод
Нейронные сети начинающим
Психология ИИ
Реализация ИИ
Реализация нейросетей
Создание беспилотных авто
Трезво про ИИ
Философия ИИ
Генетические алгоритмы
Капсульные нейросети
Основы нейронных сетей
Промпты. Генеративные запросы
Распознавание лиц
Распознавание образов
Распознавание речи
Творчество ИИ
Техническое зрение
Чат-боты
Авторизация
2018-10-04 11:32

Половое размножение позволяет перемешивать родительские геномы и избавляться от вредных мутаций. Появление секса ускорило эволюцию, потому что под действие отбора стали попадать не отдельные особи, а конкретные гены. Это дало такое преимущество, что организмов, размножающихся исключительно бесполым путем, то есть не имеющих рекомбинации и межорганизменного перемешивания генов, осталось не так уж много.
Бделлоидные коловратки: исключение из правила
Если мы посмотрим на распределение на эволюционном древе форм, отказавшихся от полового размножения, мы часто увидим концевые веточки, которые возникли недавно. Старые бесполые ветви почти отсутствуют, за исключением бделлоидных коловраток. Это единственный класс животных, который отказался от полового размножения почти 100 миллионов лет назад и до сих пор не вымер.
Одно из важных преимуществ, которое дает половое размножение, — сопротивление к паразитам. Если организм клонально размножается и все потомство генетически одинаково, паразиты очень быстро научатся эффективно бороться с защитой, а этот клон будет под угрозой. Но если гены рекомбинируют и появляются варианты и комбинации в том числе защитных генов, паразитам гораздо сложнее адаптироваться.
Бделлоидные коловратки необычны во многих отношениях. Во-первых, они умеют выдерживать полное высыхание на любой жизненной стадии, в том числе и во взрослом состоянии. Если пересыхает лужа, где они живут, их может подхватить ветром и перенести в другой водоем. Коловратки размножаются клонально, но периодически избавляются от паразитов за счет того, что они высыхают полностью и переносятся в другой водоем. Такую экзекуцию мало кто выдержит, поэтому коловратки успешно избавляются от своих паразитов, не прибегая к генетическим манипуляциям.
Бделлоидная коловратка Adenita sp© George Barron, atrium.lib.uoguelph.ca
Во-вторых, только у бделлоидных коловраток, единственных из всех животных, обнаружено множество чужих генов в геноме. Предполагается, что они занимаются горизонтальным переносом, когда высыхают: клеточные мембраны рвутся, хромосомы тоже рвутся. Когда коловратки размокнут и начнут оживать, возникнет необходимость в репарации разрывов хромосом, а через трещинки может проникнуть чужая ДНК — водорослей, бактерий, грибов, других коловраток. И при репарации двойных разрывов с определенной вероятностью происходит встраивание чужих кусочков.
Ученые давно хотели проверить, нет ли у этих коловраток также и внутривидового горизонтального переноса. Я слышал от коллег, что такое исследование проведено и вроде бы внутривидовой перенос подтвердился, но работа, насколько мне известно, еще не опубликована. Таким образом, полового размножения у бделлоидных коловраток нет, но вместо этого есть горизонтальный перенос и умение убегать от паразитов экзотическим способом.
Все остальные животные, утратившие половое размножение, в эволюционном масштабе времени быстро вымирают. При секвенировании геномов микробных популяций оценка интенсивности горизонтального переноса обычно очень высока. Хотя и считается, что бактерии размножаются клонально, на самом деле у них есть механизм перемешивания геномов внутри популяции. При клональном размножении все гены должны наследоваться сцепленно, но у бактерий де-факто локусы не сцеплены: если два гена встречаются каждый в двух вариантах, то в этой микробной популяции будут встречаться не три комбинации из четырех возможных, как следовало бы ожидать при клональном размножении, а все четыре, что указывает на горизонтальный перенос.
Локус – местоположение определённого гена на карте хромосомы // bio3400.nicerweb.net
Что считать бесполым размножением
Если бесполым размножением называть все способы получения потомства от одного родителя, можно выделить три основных варианта. Клональное размножение — это размножение без всякой рекомбинации, простое создание клонов родителя. Некоторые черви, как и микроорганизмы, практикуют деление пополам или почкование, растения — вегетативное размножение частями корневища или стебля.
Второй способ бесполого размножения — партеногенез, но он тоже бывает очень разным. Диплоидная самка может откладывать диплоидное яйцо, и тогда потомки — это ее клоны. У пчел и перепончатокрылых самки диплоидные, у них происходит мейоз. Самка откладывает гаплоидное яйцо с половиной своих генов, и из него развивается самец. Самцы гаплоидные, и, если они оплодотворяют какие-то яйца, получаются снова диплоидные самки.
Наконец, может происходить самооплодотворение, как у нематоды С. elegans: она производит спермий и яйцеклетку, которые оплодотворяют друг друга. И хотя родитель один, какая-то рекомбинация на уровне материнских гамет все же произошла. Толку от этого мало, но потомство не будет генетической копией родителя, если родитель был гетерозиготным по каким-то локусам.
Почему нужно два пола
Размножение и рекомбинация генов необязательно должны быть слиты в единый процесс. Но в любом случае секс предполагает встречу с каким-то другим организмом. Мы привыкли к тому, что полов два, но есть виды, у которых не два пола, а множество типов спаривания. Их могут быть десятки, но главная идея в том, что при такой схеме размножения организм не может скрещиваться со своим типом спаривания, а может с любым другим. Так размножаются многие амебы, в целом у низших эукариот такое встречается довольно часто. Но в непосредственном обмене генетическим материалом участвуют всегда только двое.
Бактерии тоже конъюгируют по двое, хотя теоретически возможно образование сетей, когда прокариоты создают цитоплазматические мостики, по которым передается ДНК, сразу между несколькими клетками. У Айзека Азимова есть замечательный роман «Сами боги» («The Gods Themselves»). Там изображена инопланетная жизнь с тремя полами, которые сливаются по трое, чтобы размножиться. Но это фантастика, неприменимая для земной жизни.
На Земле в основе идеи полового размножения лежит гомологичная рекомбинация: две похожие молекулы ДНК могут поменяться участками. Раз в рекомбинации участвуют две молекулы ДНК, значит, возможно только два партнера. Наверное, движущую силу полового размножения в конечном счете можно свести к митозу и его механизмам. Митоз — это бинарное деление. Каждая хромосома удваивается, а не утраивается, а потом каждая хроматида идет в дочернюю клетку. Клетка делится на две, а не на три.
Мейоз тоже развился на основе митоза. И поскольку изначально размножение было основано на бинарном делении, то уже к имеющимся механизмам подстраивались и мейоз, и сингамия. Если мы три клетки сольем вместе, нет готового механизма, чтобы обратно на три разделить, но есть готовый механизм, чтобы разделить на две. Поэтому для секса нужны двое.
Горизонтальный перенос появился, скорее всего, еще до клеток, когда существовал мир РНК. В экспериментах воспроизводится рекомбинация между молекулами РНК, затем мог возникнуть слабо упорядоченный, диффузный обмен генетическим материалом между молекулами ДНК, потом между клетками у прокариот. А митоз и мейоз — это уже ноу-хау эукариот. Эти процессы появляются вместе с эукариотической клеткой, ни раньше, ни позже.
Бактерии тоже делятся, но не митозом. У них совсем другие механизмы. Например, единственная кольцевая хромосома удваивается, две копии приклеиваются к разным местам клеточной мембраны, а затем, когда клетка делится, два колечка оказываются в разных дочерних клетках. Митоз — это специфический механизм, который позволяет точно по копиям распределить между двумя клетками много хромосом после репликации.

Поделиться![]()
![]()
![]()
![]()
Схема митоза: 1) Интерфаза 2) Профаза 3) Прометафаза 4) Метафаза 5) Анафаза 6) Телофаза и цитокинезwikipedia.org
Бывают полиплоидные прокариоты, у которых много хромосом в каждой клетке, но они распределяются по дочерним клеткам случайным образом. Генетика такого деления очень необычна: возникают странные эволюционно-генетические последствия. Например, поскольку нет гарантий, что каждая дочерняя клетка получит ровно по одной копии каждой хромосомы, множественные хромосомы не могут поделить функции. Остается простой полиплоидный организм: если ему нужно зачем-то, чтобы у него было 20 копий одной хромосомы в клетке, так будет, и они останутся одинаковыми до тех пор, пока не возникнет митоз.
Секс и эволюция человека
Возникновение полового размножения со страшной силой должно ускорять эволюцию. Хотя накопление вредных мутаций замедляется, накопление полезных мутаций должно сильно ускоряться. Адаптация к новым условиям тоже ускоряется за счет полового размножения. Это можно проверить экспериментально: сравнить две популяции дрожжей, бесполую и половую, и оценить, насколько хорошо они адаптируются к новым условиям. Это можно определить по количеству потомков в единицу времени или по скорости роста культуры по сравнению с неэволюционировавшим предком, который лежит в холодильнике. В ходе подобных экспериментов было показано, что у бесполых дрожжей приспособленность за тысячу поколений вырастает на 8%, а у половых — на 18%.
Скорость адаптации зависит от многих факторов, например от численности популяции. Преимущества полового размножения будут гораздо заметнее в большой популяции, чем в маленькой, исходя из механизма Фишера — Мёллера и эффекта накопления полезных мутаций. В большой популяции возникает больше разных мутаций, а клональное размножение не позволяет комбинировать полезные мутации. Но в маленькой популяции их и мало возникает, поэтому комбинировать особо нечего, и преимущества полового размножения будут не так заметны. В большой популяции возникает очень много разных полезных мутаций и есть что комбинировать. Поэтому половое размножение дает преимущество: в огромной бесполой популяции весь этот миллион полезных мутаций сохраниться не может, и они будут только конкурировать и вытеснять друг друга.
Секс делает все полезные мутации популяции «общим достоянием». Поэтому скорость приспособления к меняющимся условиям у организмов, способных к сексу, должна быть выше, чем у бесполых. Эту идею первыми разработали в 1930-х годах генетик Герман Мёллер и эволюционист Рональд Фишер.
Человеческая популяция стала огромной, с развитием транспорта и глобализации начали быстрее перемешиваться генофонды и размываться генетические и фенотипические границы между популяциями. Становится все больше смешанных браков, растет гетерозиготность потомства. В последние годы очень быстро снижается средний уровень родства родителей. Недавно в Science была опубликована интересная статья: исследователи обработали огромные массивы данных с сайтов, где люди восстанавливают свою родословную. Эти данные образуют гигантские сети с миллионами людей, и это уже можно обрабатывать статистически.
Оказалось, что среднее расстояние между местами рождения мужа и жены увеличилось в десятки раз. Если триста лет назад это было порядка нескольких километров, то есть люди женились на ком-то из своей деревни или в крайнем случае из соседней, то с момента появления нормального транспорта, кораблей и железных дорог это значение пошло вверх. Сегодня среднее расстояние между местом рождения мужа и жены сотни километров. Аналогичный график можно построить и по средней степени родства мужа и жены. Триста лет назад это было довольно близкое родство, на уровне четвероюродных и пятиюродных братьев и сестер. А сейчас это в два-три раза меньше, то есть браки стали менее близкородственными.
С одной стороны, это должно повышать гетерозиготность потомства и генетическое разнообразие популяции. Это в целом хорошо с точки зрения эволюционной эпидемиологии и отчасти с точки зрения здоровья потомства: такая перемешиваемость близка к панмиксии. С другой стороны, исчезновение популяционной структуры вида и гомогенизация генофонда чисто теоретически могли бы иметь и негативный эффект, но поскольку эта популяция расселена по всему земному шару и все равно разные ее части живут в очень разных условиях, то, скорее всего, эти плохие эффекты не проявятся.
Забота о потомстве
Считается, что высокий генетический полиморфизм и высокая гетерозиготность повышают шансы популяции на выживание. И оказалось, что генетический полиморфизм выше у тех, кто не заботится о потомстве, — у r-стратегов, которые мечут миллион икринок. Казалось бы, с точки зрения генетики это очень выгодная стратегия.
Когда вы вкладываете много ресурсов в каждого потомка, вместо миллиона икринок вы выращиваете трех детенышей, но зато заботитесь о них, выкармливаете и выпускаете в жизнь уже большими, сильными, умными, упитанными, и они все с большой вероятностью выживут. Генетический полиморфизм у таких видов оказался намного меньше, но тем не менее K-стратеги не вымирают. В долгосрочной перспективе они, наоборот, выигрывают. И если в отношении морской биоты можно еще спорить, потому что там не все так очевидно, то наземная биота идет в сторону K-стратегии — заботы о потомстве (тенденции можно проследить от первых мхов и амфибий).
Растения тоже стали больше вкладываться в потомство: сначала были споровые, потом голосеменные и, наконец, покрытосеменные растения, у которых семена защищены и снабжены запасом питательных веществ. А начиналось все с плавающих в лужах гамет. У тетрапод то же самое: сравните амфибий, которые мечут икру, и млекопитающих, которые вкладывают огромное количество ресурсов в каждого потомка. Во время экологических кризисов и массовых вымираний исчезают больше r-стратеги. Динозавры плохо заботились о потомстве и вымерли. Млекопитающие хорошо заботились — выжили. Аммониты, у которых были мелкие потомки, вымерли. Живое ископаемое наутилус выжило, а из яйца у него выходят уже достаточно крупные животные.

Поделиться![]()
![]()
![]()
![]()
Nautilus pompilius© wikipedia.org
Видимо, не все так плохо у K-стратегов: можно жить и с низким полиморфизмом. Если рожать много детенышей с высокой смертностью, это дает много возможностей для отбора. Если из тысячи выживает пять, наверное, это значит, что отбор хороший: все плохие гены отбраковались. А если вы рожаете пять детенышей и из них выживает четыре, то какой тут, казалось бы, отбор. Будут накапливаться вредные мутации, и популяция вымрет.
Я думаю, что, когда у r-стратегов из тысячи выживает четыре, это случайная смертность, мало зависящая от качества генов. Все решается по принципу «повезло — не повезло». Если кит плывет и фильтрует криль, он просто входит в стаю и засасывает всех подряд. Кого он засосет, а кого не засосет — совершенно не зависит от качества генов конкретного рачка. Это случайная смертность, хоть и высокая. Другое дело, если есть мощная забота о потомстве и упитанный детеныш выходит в жизнь. Родители в него вложили все, элемент случайности в его дальнейшей судьбе сведен к минимуму. Дальше его успех зависит уже от его личных качеств, в том числе от качества генов. И хотя смертность ниже, отбор, возможно, получается более эффективным.
Теория r/K-отбора — экологическая теория, определяющая две стратегии размножения живых организмов. Разные стратегии обеспечивают оптимальную численность для разных видов. Теорию разработали два американских эколога — Роберт МакАртур и Эдвард Уилсон. Впервые она была опубликована в 1967 году в работе «Теория островной биогеографии»
Как проводить границы между видами
Границы между видами размыты. Обычно в реальной палеонтологической летописи какие-то границы мы наблюдаем. Они связаны, как правило, с неполнотой летописи. Если между двумя ископаемыми формами перерыв составляет 2 миллиона лет, то мы видим разные виды. А если бы у нас была полная летопись, мы бы видели постепенный переход. Конечно, изменения идут не с постоянной скоростью: изменились условия — быстро пошел отбор. Но все равно это не мгновенно.
Еще Дарвин прекрасно понимал эту ситуацию, и он написал следующее в книге «Происхождение человека»: в бесконечном ряду форм, постепенно переходящих от какого-то обезьяноподобного предка к человеку в его современном состоянии, невозможно было бы точно указать ту форму, которой впервые следует присвоить имя человека. Поэтому в таких случаях принимают какое-то чисто формальное решение: давайте всех, у кого мозг меньше 600 кубических сантиметров, считать австралопитеками, а тех, у кого больше, — Homo. Такой подход тоже не срабатывает, потому что даже у представителей одной популяции очень большая изменчивость. Пять черепов из Дманиси, которые почти наверняка представляют одну популяцию, — это ранние Homo erectus, и у них такая изменчивость по размеру мозга, что, если формально применять все критерии, из родных братьев можно отнести одного к обезьянам, а другого — к людям.

![]()
![]()
Череп из Дманиси D2700© wikipedia.org
В современной фауне мы видим много четких видов — просто потому, что переходные формы вымерли. Но также мы видим и случаи видообразования, которые происходят прямо на наших глазах. И тогда действительно непонятно, считать эти формы разными видами, разновидностями, подвидами или расами. Есть некая традиция, устоявшийся свод условных правил, по которым зоологи решают, когда мы считаем две формы видами, а когда подвидами или разновидностями. Принято считать, что если между двумя формами в природе скрещиваний не бывает или почти никогда не бывает и они при этом более-менее отличаются друг от друга генетически, фенотипически или по поведению, то это разные виды. Но это все очень зыбко.
Например, белый медведь и бурый медведь — это два типичных вида. Ни один зоолог не скажет, что это один и тот же вид: Ursus maritimus — белый медведь, Ursus arctos — бурый медведь, или гризли, как его называют в Америке. Давно было известно, что в зоопарках они прекрасно скрещиваются и дают плодовитое гибридное потомство, но в природе они не гибридизируются, потому что не встречаются.
В последние десятилетия климат стал меняться, арктические льды тают. Белые медведи не могут найти себе подходящих льдин и вылезают на берег, встречаются там с гризли и иногда скрещиваются, то есть уже в природе они стали давать гибридное потомство. Первый раз это обнаружили, когда охотник на Аляске купил лицензию на гризли, застрелил медведя, а лесники ему говорят: «Что ты сделал? Ты белого медведя застрелил». Стали спорить, провели генетический анализ, и оказалось, что это гибрид. Потом еще где-то в Гренландии нашли целую группу гибридных медведей. Оказалось, одной белой самке нравилось спариваться с бурыми самцами. Она нарожала гибридного потомства, которое тоже начало с кем попало скрещиваться.
Считать ли их теперь одним видом? В принципе, есть шанс, что они могут полностью смешаться. Если так пойдет и дальше, совсем не останется мест, где белые медведи смогут по-своему жить, и им придется возвращаться на сушу. Они приспособлены жить на льдинах и охотиться на тюленей, а если льдин не будет, они могут влиться обратно в генофонд бурых медведей. И таких ситуаций полно, это недавнее видообразование. Несколько сотен тысяч лет назад виды разделились, и в прошлом были эпизоды гибридизации.
Аналогичная ситуация была, похоже, с сапиенсами, неандертальцами и денисовцами. С одной стороны, они гибридизировались, при встрече скрещивались друг с другом, давали потомство, небольшие порции чужеродных генов проникали в генофонды. У денисовцев и сапиенсов есть небольшая примесь неандертальцев, а вот у поздних европейских неандертальцев не нашли примеси сапиенса. Показали, что у них была довольно значительная постзиготическая изоляция, то есть, по-видимому, пониженная приспособленность гибридов. Значит, была частичная генетическая несовместимость. Поэтому чужие гены могли проникнуть в генофонд другого вида, но многие из них оказывались вредными и постепенно вычищались отбором. Большая часть неандертальской примеси, которую мы получили, с тех пор постепенно вычищается отбором. Если существует генетическая несовместимость, пусть и неполная, это довод в пользу того, чтобы считать разными эти два вида.
Есть много примеров межвидовой гибридизации: разные виды собак, все шакалы, койоты, волки свободно скрещиваются друг с другом в любых комбинациях, если у них есть такая возможность. Никакой четкой границы между видами и разновидностями нет.
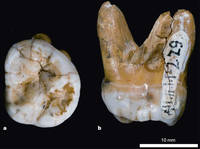
Денисовский человек — предположительный вид или подвид вымерших людей, известный по фрагментарному материалу пяти находок из Денисовой пещеры в Солонешенском районе Алтайского края России. На рисунке изображен знаменитый зуб из Денисовой пещеры //David Reich et al., Nature, 2010
Поиск общих предков
У любых двух современных видов когда-то был общий предок. Этого общего предка можно назвать переходной формой. Настоящие переходные формы не могут жить одновременно с двумя современными видами, потому что они жили в прошлом. Современные организмы на то и современные, что никто из них не произошел ни от кого другого. У них были общие предки, но шимпанзе, например, не произошел от человека, а человек не произошел от шимпанзе. По каким-то признакам шимпанзе меньше изменился по сравнению с общим предком, чем человек. Например, у человека вырос мозг в три раза, а у шимпанзе не вырос. По размеру мозга можно тогда сказать, что шимпанзе является переходной формой между мартышкой и человеком, но все относительно, если мы рассматриваем отдельные признаки.
Определить, сколько времени прошло между расхождением двух видов, можно, пользуясь гипотезой молекулярных часов. Технически это довольно сложно, потому что скорость мутагенеза все же неодинаковая, поэтому молекулярные часы идут не очень точно. Либо нужно доказывать, что у изучаемых видов скорость мутагенеза одинаковая, либо, если она разная, нужно вносить поправки на эти различия и калибровать молекулярные часы на основе того, что мы знаем. Сейчас, когда строятся филогенетические деревья с использованием молекулярных часов, по умолчанию принимается, что эти молекулярные часы в разных ветвях могут идти с разной скоростью.
Поэтому стараются откалибровать дерево по максимальному количеству ископаемых датированных находок и таким образом ограничить во времени определенные узлы дерева. Если есть ископаемые змеи и ящерицы среднеюрского или позднемелового возраста, значит, разделение змей и ящериц произошло раньше. Зная по палеонтологии, когда разделились змеи и ящерицы, имея геномы змей и ящериц, мы можем оценить скорость накопления различий у змей и ящериц. И тогда, если у нас есть еще пять разных змей, оценив эту скорость, мы можем оценить и время расхождения пяти разных видов. Но для каждой веточки будет своя опорная точка и свои расчеты.
Можно попробовать вычислить последнего общего предка всех ныне живущих людей, но обычно такой задачи не ставят, потому что результат будет смешной. Выяснится, что этот последний общий предок жил на удивление недавно, потому что здесь геометрическая прогрессия: у каждого человека два родителя, четыре дедушки и бабушки, восемь прадедушек и прабабушек и так далее. За счет этой чудовищно быстро нарастающей прогрессии получается, что достаточно одного индуса на пироге, случайно занесенного в Австралию тысячу лет назад, чтобы он оказался предком всех аборигенов, хотя понятно, что ДНК конкретно этого индуса у большинства нет или почти нет.
Исходя из этого, я совершенно уверен, что можно почти строго доказать: вы или я потомок Чингисхана, потому что у таких людей, как он, было много детей, много внуков, есть кое-какие оценки, за которые можно зацепиться, и, учитывая эту геометрическую прогрессию, современное число потомков таких людей, как Чингисхан или Рюрик, на много порядков превышает население Земли, просто если рассчитать эту геометрическую прогрессию.
Поэтому последний общий предок всех людей жил недавно. А вот последний общий предок всех людей чисто по женской линии жил намного раньше — 150 тысяч лет назад, в Африке. Ее называют митохондриальная Ева. Y-хромосомный Адам тоже жил более сотни тысяч лет назад. Но когда в Африке находят какую-нибудь неизвестную ранее гаплогруппу Y-хромосом, это сразу может удревнить Адама. Но это не значит, что митохондриальная Ева или Y-хромосомный Адам были какими-то особо важными узловыми точками в эволюции человечества. Это просто статистический фокус: некая реальная женщина и некий реальный мужчина, которым случайно повезло оказаться последними общими предками всех современных людей по чисто мужской или по чисто женской линии. Ничего особо осмысленного в этом нет.
Оценку можно проводить по межпопуляционным различиям и времени расхождения популяций. Это имеет какой-то смысл, если мы знаем скорость мутагенеза. Согласно последним оценкам, предки людей и шимпанзе разделились 13 миллионов лет назад, хотя раньше говорили, что 7. Стоило более точно померить скорость мутагенеза, и она оказалась меньше, чем ожидали, — пришлось все удревнять. Сейчас для повышения точности метода секвенируют тройки «мама — папа — ребенок» и буквально вручную считают новые мутации, которые есть у ребенка. Это сделали и для людей, и для шимпанзе.
Все палеоантропологи уже успели привыкнуть к 7 миллионам лет, а теперь все сильно меняется. Значит, оррорин и сахелантроп, которые считались формами, очень близкими к последнему общему предку человека и шимпанзе, оказываются очень далекими формами, то есть жившими намного позже. Значит, становится актуальным вопрос о том, к какой линии они принадлежат: к нашей, к линии шимпанзе или к какой-то третьей, вымершей. По морфологии они как раз очень похожи на имеющиеся представления об общем предке.

Поделиться![]()
![]()
![]()
![]()
Модель головы Sahelanthropus tchadensis в музее Естественной истории, США© flickr.com
Внешний вид ископаемых форм восстанавливается по принципу актуализма: когда мы находим кости какого-нибудь грызуна, то легко его восстановить, потому что мы знаем, как устроены грызуны. А если мы находим представителя какого-то вымершего типа, возникают проблемы. Есть находки, которые неизвестно как интерпретировать, и приходится искать уникальные экземпляры с уникальной сохранностью. Но все равно есть группы ископаемых, которые так и остаются в статусе неизвестных.
Мощное оружие — это анализ ДНК, но если ископаемому 500 миллионов лет, то там никакой ДНК нет. К тому же фенотип зависит не только от генов. Все признаки развиваются на основе сложных взаимодействий генов и среды. С одним и тем же геномом в зависимости от питания вы можете быть разного размера. Конечно, научиться по генотипу предсказывать фенотип с максимально возможной точностью — это великая задача. Но сначала надо очень хорошо разобраться в том, как устроен онтогенез, а до этого еще тоже далеко.
Вопросов пока больше, чем ответов, как всегда бывает в науке. Хотя сегодня мы знаем про эволюцию человека настолько больше, чем, скажем, пятнадцать лет назад, что это потрясает, прогресс очень быстрый. Но опять же, чем быстрее прогресс в науке, тем быстрее возникают и новые вопросы. Мы работаем с дрозофилами — казалось бы, плодовая мушка, про нее все известно, каждый ген. Ее изучают сто лет десятки тысяч ученых. А такое впечатление, что конца и края нет — ничего не известно про дрозофилу.
Телеграм: t.me/ainewsline
Источник: postnauka.ru