Таламус
МЕНЮ
Главная страница
Поиск
Регистрация на сайте
Помощь проекту
Архив новостей
ТЕМЫ
Новости ИИ
Городские сумасшедшие
ИИ в медицине
ИИ проекты
Искусственные нейросети
Искусственный интеллект
Слежка за людьми
Угроза ИИ
ИИ теория
Компьютерные науки
Машинное обуч. (Ошибки)
Машинное обучение
Машинный перевод
Нейронные сети начинающим
Психология ИИ
Реализация ИИ
Реализация нейросетей
Создание беспилотных авто
Трезво про ИИ
Философия ИИ
Генетические алгоритмы
Капсульные нейросети
Основы нейронных сетей
Промпты. Генеративные запросы
Распознавание лиц
Распознавание образов
Распознавание речи
Творчество ИИ
Техническое зрение
Чат-боты
Авторизация
2018-04-28 08:00


Таламус
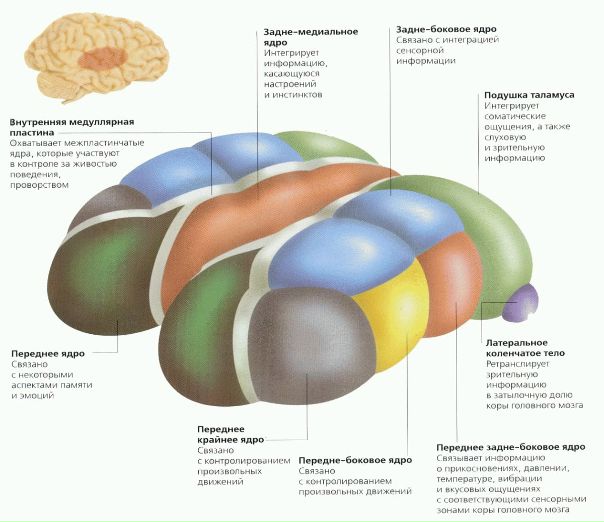
Таламус (thalamus, зрительный бугор) — структура, в которой происходит обработка и интеграция практически всех сигналов, идущих в кору большого мозга от спинного, среднего мозга, мозжечка, базальных ганглиев головного мозга.
Морфофункциональная организация. В ядрах таламуса происходит переключение информации, поступающей от экстеро-, проприорецепторов и интероцепторов и начинаются таламокортикальные пути.
Учитывая, что коленчатые тела таламуса являются подкорковыми центрами зрения и слуха, а узел уздечки и переднее зрительное ядро участвуют в анализе обонятельных сигналов, можно утверждать, что зрительный бугор в целом является подкорковой «станцией» для всех видов чувствительности. Здесь раздражения внешней и внутренней среды интегрируются, после чего поступают в кору большого мозга.
Зрительный бугор является центром организации и реализации инстинктов, влечений, эмоций. Возможность получать информацию о состоянии множества систем организма позволяет таламусу участвовать в регуляции и определении функционального состояния организма в целом (подтверждением тому служит наличие в таламусе около 120 разнофункциональных ядер). Ядра образуют своеобразные комплексы, которые можно разделить по признаку проекции в кору на 3 группы: передняя проецирует аксоны своих нейронов в поясную извилину коры большого мозга; медиальная — в лобную долю коры; латеральная — в теменную, височную, затылочную доли коры. По проекциям определяется и функция ядер. Такое деление не абсолютно, так как одна часть волокон от ядер таламуса идет в строго ограниченные корковые образования, другая — в разные области коры большого мозга.
Ядра таламуса функционально по характеру входящих и выходящих из них путей делятся на специфические, неспецифические и ассоциативные.
К специфическим ядрам относятся переднее вентральное, медиальное, вентролатеральиое, постлатеральное, постмедиальное, латеральное и медиальное коленчатые тела. Последние относятся к подкорковым центрам зрения и слуха соответственно.
Основной функциональной единицей специфических таламических ядер являются «релейные» нейроны, у которых мало дендритов и длинный аксон; их функция заключается в переключении информации, идущей в кору большого мозга от кожных, мышечных и других рецепторов.
От специфических ядер информация о характере сенсорных стимулов поступает в строго определенные участки III—IV слоев коры большого мозга (соматотопическая локализация). Нарушение функции специфических ядер приводит к выпадению конкретных видов чувствительности, так как ядра таламуса, как и кора большого мозга, имеют соматотопическую локализацию. Отдельные нейроны специфических ядер таламуса возбуждаются рецепторами только своего типа. К специфическим ядрам таламуса идут сигналы от рецепторов кожи, глаз, уха, мышечной системы. Сюда же конвергируют сигналы от интерорецепторов зон проекции блуждающего и чревного нервов, гипоталамуса.
Латеральное коленчатое тело имеет прямые эфферентные связи с затылочной долей коры большого мозга и афферентные связи с сетчаткой глаза и с передними буграми четверохолмий. Нейроны латеральных коленчатых тел по-разному реагируют на цветовые раздражения, включение, выключение света, т. е. могут выполнять детекторную функцию.
В медиальное коленчатое тело (МТК) поступают афферентные импульсы из латеральной петли и от нижних бугров четверохолмий. Эфферетные пути от медиальных коленчатых тел идут в височную зону коры большого мозга, достигая там первичной слуховой области коры. МКТ имеет четкую тонотопичность. Следовательно, уже на уровне таламуса обеспечивается пространственное распределение чувствительности всех сенсорных систем организма, в том числе сенсорных посылок от интерорецепторов сосудов, органов брюшной, грудной полостей.
Ассоциативные ядра таламуса представлены передним медиодорсальным, латеральным дорсальным ядрами и подушкой. Переднее ядро связано с лимбической корой (поясной извилиной), медиодорсальное — с лобной долей коры, латеральное дорсальное — с теменной, подушка — с ассоциативными зонами теменной и височной долями коры большого мозга.
Основными клеточными структурами этих ядер являются мультиполярные, биполярные трехотростчатые нейроны, т. е. нейроны, способные выполнять полисенсорные функции. Ряд нейронов изменяет активность только при одновременном комплексном раздражении. На полисенсорных нейронах происходит конвергенция возбуждений разных модальностей, формируется интегрированный сигнал, который затем передается в ассоциативную кору мозга. Нейроны подушки связаны главным образом с ассоциативными зонами теменной и височной долей коры большого мозга, нейроны латерального ядра — с теменной, нейроны медиального ядра — с лобной долей коры большого мозга.
Неспецифические ядра таламуса представлены срединным центром, парацентральным ядром, центральным медиальным и латеральным, субмедиальным, вентральным передним, парафасцикулярным комплексами, ретикулярным ядром, перивентрикулярной и центральной серой массой. Нейроны этих ядер образуют свои связи по ретикулярному типу. Их аксоны поднимаются в кору большого мозга и контактируют со всеми ее слоями, образуя не локальные, а диффузные связи. К неспецифическим ядрам поступают связи из РФ ствола мозга, гипоталамуса, лимбической системы, базальных ганглиев, специфических ядер таламуса.
Возбуждение неспецифических ядер вызывает генерацию в коре специфической веретенообразной электрической активности, свидетельствующей о развитии сонного состояния. Нарушение функции неспецифических ядер затрудняет появление веретенообразной активности, т. е. развитие сонного состояния.
Сложное строение таламуса, наличие в нем взаимосвязанных специфических, неспецифических и ассоциативных ядер, позволяет ему организовывать такие двигательные реакции, как сосание, жевание, глотание, смех. Двигательные реакции интегрируются в таламусе с вегетативными процессами, обеспечивающими эти движения.
Конвергенцию сенсорных стимулов в таламус обусловливает возникновение так называемых таламических неукротимых болей, которые возникают при патологических процессах в самом таламусе.
Телеграм: t.me/ainewsline
Источник: vk.com