Главы | Начало секса
МЕНЮ
Главная страница
Поиск
Регистрация на сайте
Помощь проекту
Архив новостей
ТЕМЫ
Новости ИИ
Городские сумасшедшие
ИИ в медицине
ИИ проекты
Искусственные нейросети
Искусственный интеллект
Слежка за людьми
Угроза ИИ
ИИ теория
Компьютерные науки
Машинное обуч. (Ошибки)
Машинное обучение
Машинный перевод
Нейронные сети начинающим
Психология ИИ
Реализация ИИ
Реализация нейросетей
Создание беспилотных авто
Трезво про ИИ
Философия ИИ
Генетические алгоритмы
Капсульные нейросети
Основы нейронных сетей
Промпты. Генеративные запросы
Распознавание лиц
Распознавание образов
Распознавание речи
Творчество ИИ
Техническое зрение
Чат-боты
Авторизация
2018-03-07 21:00

Отрывок из книги британского биохимика Ника Лейна, посвященный половому процессу, рекомбинации генов и естественному отбору
Совместно с издательством Corpus мы публикуем отрывок из книги британского биохимика Ника Лейна «Вопрос жизни», посвященной эволюции, происхождению сложности и процессам, лежащим в основе жизни и смерти. Перевод с английского Ксении Сайфулиной и Матвея Колесника.
Настоящий секс, который практикуют эукариоты, предполагает слияние двух гамет (в нашем случае сперматозоида и яйцеклетки) с половиной набора хромосом у каждой. Мы, как и многие другие многоклеточные эукариоты, диплоидны: в наших клетках каждый ген присутствует в двух экземплярах — от отца и от матери. Если точнее, у нас по два экземпляра каждой хромосомы. На картинках в учебниках хромосомы выглядят так, будто это стабильные физические структуры, но это далеко не так. В ходе образования гамет хромосомы рекомбинируют — обмениваются друг с другом участками. Благодаря этому появляются новые комбинации генов, которых, скорее всего, прежде не бывало.
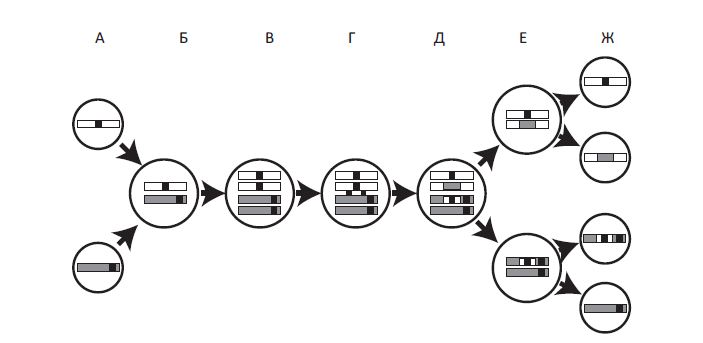
Половой процесс и рекомбинация у эукариот. Приведена упрощенная схема полового цикла: слияние двух гамет, за которым следует двухстадийный мейоз. В ходе мейоза осуществляется рекомбинация, что позволяет получить новые, генетически различные гаметы. Две гаметы, содержащие по одной хромосоме из гомологичной пары (хромосомы различаются генетически), сливаются (А). Гаметы образуют зиготу с двойным набором хромосом (Б). (Черные полоски могут обозначать и вредную мутацию, и полезный вариант какого-либо гена.) Перед мейозом хромосомы удваиваются (В). После этого в клетке образуется по две эквивалентных копии каждой хромосомы (каждая хромосома теперь состоит из двух сестринских хроматид). Затем две или более хромосомы попарно рекомбинируют (Г). Хромосомы совершают взаимообмен участками ДНК, и на выходе мы получаем обновленные хромосомы, каждая из которых включает фрагменты обеих исходных: и материнской, и отцовской (Д). При первом делении мейоза хромосомы разделяются, попадая в две разные клетки (Е), а в ходе второго деления образуются гаметы (Ж). Две из получившихся гамет эквивалентны исходным, две другие — нет. Если черные полоски обозначают вредные мутации, то в результате полового процесса образовалась одна гамета без мутаций и одна с двумя мутациями. Последняя будет элиминирована в ходе естественного отбора. Если черные полосы обозначают полезные мутации, то половой процесс объединил их в одной гамете, давая возможность естественному отбору воздействовать на них одновременно. Короче, половое размножение приводит к повышению многообразия гамет, делая их «заметнее» для отбора, и способствует элиминации вредных мутаций и распространению полезных.
Если вы пройдетесь по только что рекомбинировавшей хромосоме, перебирая ген за геном, то обнаружите, что она включает в себя гены обоих родителей. В ходе мейоза (редукционного деления) две хромосомы разделяются и попадают в гаплоидные гаметы, в которых каждая хромосома должна быть представлена одной копией. В итоге две гаметы (каждая содержит рекомбинантные хромосомы) сливаются — и получается оплодотворенная яйцеклетка, индивид с уникальной комбинацией генов, будущий ребенок. Проблема с возникновением полового процесса состоит не в том, что для этого требовалось выработать множество новых механизмов. В ходе рекомбинации две гомологичные хромосомы конъюгируют, плотно прикрепляясь друг к другу. Затем хромосомы крест-накрест обмениваются некоторыми участками. Конъюгация хромосом и генетическая рекомбинация также осуществляются бактериями и археями в ходе горизонтального переноса генов, но она не всегда взаимна и применяется для того, чтобы починить поврежденные хромосомы или вновь обрести утерянные гены. Применяемые здесь молекулярные механизмы в основном те же. Рекомбинация при половом процессе отличается более широкими масштабами, а также тем, что обмен всегда взаимен. Половое размножение — это взаимная рекомбинация, происходящая по всей длине генома. Если такое и встречается у прокариот, то очень редко.
В XX веке проблема смысла полового размножения была настоящей «царицей» биологических проблем, но сейчас мы уже понимаем, чем оно полезно: хотя бы тем, что пользоваться половым процессом лучше, чем размножаться исключительно бесполым путем (клонированием). Половой процесс разбивает устойчивые сочетания генов, позволяя естественному отбору «увидеть» гены по отдельности, перебрать по одному все наши признаки. Так можно избавиться от паразитов, приспособиться к изменяющимся условиям и поддерживать необходимый уровень разнообразия в популяции. Подобно тому, как средневековые каменотесы тщательно, со всех сторон, обрабатывали скульптуры для ниш в стенах соборов (ничто не укроется от ока Господа!), так и половой процесс тщательно обрабатывает геномы, позволяя естественному отбору детально, ген за геном, оценить собственные творения. Половое размножение делает наши хромосомы «пластичными», позволяя разным вариантам генов — аллелям — комбинироваться[1 ], и благодаря этому естественный отбор выявляет даже незначительные различия между организмами.
Представьте себе сто генов, собранных в хромосоме, которая никогда не рекомбинирует. Отбор будет действовать лишь на целую хромосому, на весь набор генов сразу. Предположим, среди этих генов есть несколько критически важных: любая мутация в одном из них почти неизбежно приведет к гибели организма. Однако мутации в менее важных генах становятся почти невидимыми для отбора. Слабовредные мутации могут накапливаться в таких генах, а негативные эффекты будут компенсироваться наличием хороших аллелей нескольких важных генов. В результате качество хромосомы и приспособленность организма будут постепенно снижаться. Примерно это и происходит с Y-хромосомой у мужчин: если рекомбинации нет, значит, большая часть генов постепенно деградирует. Естественный отбор может обеспечить сохранение качества лишь критически важных генов. Целая половая хромосома может быть утеряна — так и случилось с Ellobius lutescens — одним из видов слепушонок. Когда действует положительный отбор, ситуация становится еще хуже. Представим, что редкая полезная мутация предоставляет настолько значительные преимущества, что очень широко распространяется в популяции. Организмы, унаследовавшие новую мутацию, начинают доминировать, и в итоге она фиксируется во всей популяции: все организмы остаются с одним и тем же вариантом гена. Но естественный отбор может «видеть» только целую хромосому, а это значит, что остальные 99 генов, входящие в нее, также зафиксируются в популяции. Это явление называют «генетическим автостопом»: вместе с полезным аллелем одного гена в популяции закрепляются и другие — он как бы везет на себе аллели других генов. Это-то и плохо.
Вообразите, что в популяции присутствует два или три варианта (аллеля) каждого гена и из них можно составить от 10 тыс. до 1 млн комбинаций. После фиксации все это многообразие пропадает, и популяция остается с одной-единственной комбинацией 100 генов — тех, которым выпало делить одну хромосому с полезным геном. Это катастрофическая потеря разнообразия. И, конечно, жалкие 100 генов — это грубое упрощение: у бесполых организмов тысячи генов остались лишь в одном варианте, а остальные были отброшены в ходе отбора. Такую форму отбора называют «селективным выметанием». «Эффективный» размер популяции заметно сокращается, и это ставит бесполых животных под угрозу вымирания[2 ]. И действительно: почти все бесполые растения и животные исчезли за несколько миллионов лет[3 ].
Эти два процесса — накопление слабовредных мутаций и потеря многообразия в ходе «селективного выметания» — в совокупности составляют селективную интерференцию. Если рекомбинации нет, то отбор, действующий на определенные гены, препятствует отбору других сцепленных с ними генов. Путем создания хромосом с комбинациями аллелей — «пластичных хромосом» — половой процесс позволяет отбору воздействовать на все гены по отдельности. Благодаря этому отбор видит все наши «пороки» и «добродетели» вплоть до нуклеотидов. В этом и заключается огромное преимущество полового размножения. Но у секса есть свои серьезные недостатки: недаром он долго считался самой значительной из эволюционных проблем. Половое размножение разрушает заведомо удачные комбинации аллелей, которые хорошо подходят для определенных условий, и случайным образом перемешивает гены, которые позволяли преуспеть нашим предкам. В каждом поколении набор генов вновь перетасовывается, не оставляя никаких шансов на рождение полных двойников гениальных личностей: например, второго Моцарта. Что хуже, есть еще так называемая «двойная цена секса». Когда делится бесполая клетка, она порождает две дочерние клетки. Каждая из них, в свою очередь, делится на две дочерние, и так далее. Численность популяции растет экспоненциально. Когда делится половая клетка, ее дочерние клетки должны слиться с аналогичными, чтобы сформировать новую клетку, которая породит еще две дочерних.
Выходит, размер бесполой популяции в каждом поколении удваивается, а численность популяции организмов с половым размножением остается прежней. Вместо того чтобы просто получить собственную копию, приятную во всех отношениях, в рамках полового размножения вы сталкиваетесь с проблемой поиска партнера, что сопряжено с эмоциональными (и финансовыми) затратами. Самцы не достаются просто так. Клонируйте себя — и не будет нужды в поединках на рогах, павлиньих хвостах и заполненных мужчинами залах заседаний. Можно будет навсегда забыть об ужасных заболеваниях, передающихся половым путем, например СПИДе и сифилисе, и о вирусах и «прыгающих генах», которые угрожают засорить наш геном. Почему половое размножение у эукариот распространено так широко — загадка. Можно предположить, что его преимущества компенсируют затраты лишь при определенных обстоятельствах. Это подтверждается тем, что микробы могут делиться бесполым путем в течение жизни примерно 30 поколений перед тем, как побаловать себя спонтанным сексом — обычно это происходит в условиях стресса. Но я бы сказал, что секс распространен шире, чем это оправданно. Возможно, это связано с тем, что у Последнего общего предка эукариот уже имелось половое размножение, и все его потомки унаследовали от него это свойство. Хотя многие микроорганизмы перестали регулярно пользоваться половым процессом, мало кому удалось отказаться от полового размножения и не вымереть. Как видите, цена абсолютного отказа от секса также очень высока. Это относится и к первым эукариотам: те из них, у которых никогда не было секса — так сказать, не сумевшие до него «додуматься», — были обречены на вымирание.
Но здесь мы возвращаемся к вопросу о горизонтальном переносе генов. Он аналогичен половому процессу в том отношении, что происходит генетическая рекомбинация и хромосомы становятся «пластичными». До недавних пор считалось, что бактерии — непревзойденные мастера клонирования. Их численность растет экспоненциально. Одна-единственная бактерия E. сoli, делясь каждые полчаса, за три дня способна образовать колонию, суммарная масса особей которой будет равна массе Земли (если, конечно, рост популяции не будет ничем ограничен). Кроме того, эти создания еще могут обмениваться генами при помощи горизонтального переноса, встраивая в свои хромосомы новые гены и утрачивая ставшие ненужными. Бактерии, устроившие вам кишечный грипп, могут отличаться от бактерий того же вида, живущих у вас в носу, на 30 % своего генома. Получается, бактерии наслаждаются преимуществами полового размножения (пластичностью хромосом) наряду со скоростью и простотой бесполого. Но их клетки не сливаются целиком и двух полов у них нет, поэтому многие недостатки полового размножения бактерий не затрагивают. Похоже, они взяли все лучшее от обоих миров. Так почему же ранние эукариоты перешли от горизонтального переноса генов к половому процессу? Популяционные генетики-математики Салли Отто и Ник Бэртон выделили три условия, при которых наиболее выражены преимущества полового размножения: высокая скорость мутаций, сильное давление отбора и значительное разнообразие внутри популяции. Видимо, именно с этим пришлось столкнуться древним эукариотам.
Сначала рассмотрим частоту мутаций. При бесполом размножении высокая скорость мутагенеза увеличивает темпы накопления слабовредных мутаций. Кроме того, происходит снижение многообразия за счет «селективного выметания», и селективная интерференция становится более жесткой. Скорость мутагенеза у древних эукариот, вероятно, была высокой, учитывая, что тогда происходила инвазия интронов. Насколько высокой, можно попытаться определить с помощью моделирования. Я занимаюсь этим вопросом с Эндрю Помянковски и Джезом Оуэном, который разбирается в физике и в то же время интересуется большими вопросами биологии. В данный момент Джез разрабатывает численную модель, которая позволила бы определить, в каких условиях половое размножение становится выгодным. Еще один фактор, который стоит принять во внимание — размер генома. Даже если скорость мутагенеза не изменяется (скажем, одна летальная мутация на 10 млрд «букв» ДНК), нельзя бесконечно увеличивать геном: по той или иной причине обязательно случится мутационная катастрофа. При такой скорости мутагенеза с клетками, геном которых включает менее 10 млрд нуклеотидов, все будет в порядке, а вот клетки с более длинным геномом погибнут, заполучив летальную мутацию. Приобретение митохондрий на ранних этапах эволюции эукариот должно было усугубить обе проблемы: они почти наверняка вызвали возрастание скорости мутагенеза и сделали возможным значительное (на несколько порядков) увеличение генома.
Вероятный ответ: половое размножение — единственный способ решения проблемы. Горизонтальный перенос генов теоретически может помочь избежать селективной интерференции при помощи рекомбинации, но работа Джеза показывает, что горизонтального переноса недостаточно. Чем большегеном, тем сложнее заполучить «правильный» ген путем горизонтального переноса. Это становится похоже на игру в рулетку. Единственный способ обеспечить полный набор работоспособных генов в геноме — регулярно производить рекомбинацию всего генома. Этого нельзя достичь путем горизонтального переноса. Здесь нужен «тотальный половой процесс»: лишь так можно осуществить полногеномную рекомбинацию.
Телеграм: t.me/ainewsline
Источник: postnauka.ru