Физиология сна: медленный (NREM) сон
МЕНЮ
Главная страница
Поиск
Регистрация на сайте
Помощь проекту
Архив новостей
ТЕМЫ
Новости ИИ
Городские сумасшедшие
ИИ в медицине
ИИ проекты
Искусственные нейросети
Искусственный интеллект
Слежка за людьми
Угроза ИИ
ИИ теория
Компьютерные науки
Машинное обуч. (Ошибки)
Машинное обучение
Машинный перевод
Нейронные сети начинающим
Психология ИИ
Реализация ИИ
Реализация нейросетей
Создание беспилотных авто
Трезво про ИИ
Философия ИИ
Генетические алгоритмы
Капсульные нейросети
Основы нейронных сетей
Промпты. Генеративные запросы
Распознавание лиц
Распознавание образов
Распознавание речи
Творчество ИИ
Техническое зрение
Чат-боты
Авторизация
2017-07-17 13:10







Физиология сна: медленный (NREM) сон.
“Обычные люди, засыпая, считают овец, а поклонники Матрицы – агентов Смитов”.
Неделю назад, дорогие друзья, мы сделали первый шаг в загадочный и неизведанный мир сна и его регуляции, разобрав роль орексина, как некоего настройщика или “дирижёра” в цикле сон/бодрствование — https://vk.cc/6T0Hpi. В этом посте мы разберем физиологию медленного (Non-Rapid Eye Movement - NREM; без движения глаз (БДГ)) сна. Итак, приступим.
Как и все в сомнологии, исследование системы сна без БДГ началось с Константина фон Экономо еще в 20 годах прошлого столетия. Однако, подтверждение как гистохимическое, так и нейрофизиологическое было получено только в конце 90-х годов — https://vk.cc/6T2dEd, https://vk.cc/6T1bCZ, https://vk.cc/6T1bQH, https://vk.cc/6T2jyl.
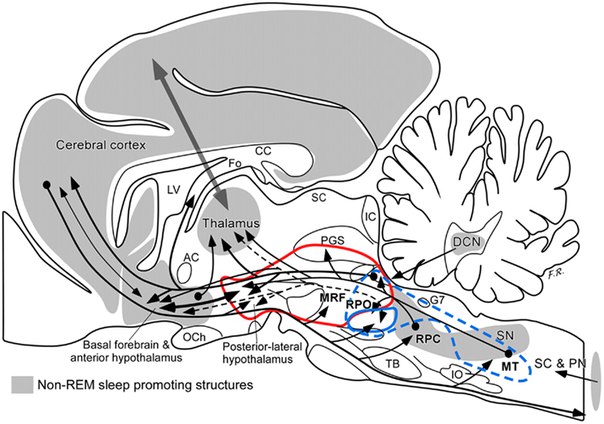
“Центр медленного сна” располагается в вентролатеральной преоптической области (ventrolateral preoptic area; VLPO) и срединной преоптической области (median preoptic area; МРО или МпРО).
Общим для данных групп нейронов является то, что они используют в качестве химического посредника одно и тоже вещество — ГАМК.
Напомню, что аксоны моноаминергической и серотонинергической систем мозга имеют окончания на ГАМК-ергических нейронах VLPO, а гистаминергическая система ТМЯ, помимо гистамина, также образует ГАМК и галанин. Орексинергические же нейроны не распространяют свое влияние на область VLPO, а вот нейроны преоптической области благодаря своим длинным аксонам тесно связаны со всеми активирующими системами мозга (через ГАМК-А и ГАМК-В рецепторы).
Таким образом, при незначительном снижении активности нейронов “системы бодрствования” происходит включение тормозных нейронов вышеописанных областей с дальнейшим снижением активности (происходит инициация и углубление сна без БДГ).
Помимо этого, в процессе углубления сна принимают участие корковые интернейроны 1-го и 2-го слоев коры и парафациальная зона, расположенная в ростральном отделе продолговатого мозга (“тормозной центр” Моруцци).
Этот процесс не длится до бесконечности, гипоталамус имеет “рубильник”, и вся система перебрасывается в другое состояние – или бодрствования, или сна с БДГ, так называемый “flip--flop” эффект. Объективно это становится заметно на ЭЭГ (1-3 фаза NREM - сна) при 90-минутном цикле.
Также в процессе дня, когда мы бодрствуем, в головном мозге происходит активное накопление “метаболитов” и медиаторов, связанных с интенсивным обменом веществ. Чем интенсивнее умственная работа, тем выше скорость метаболизма и больше образуется метаболитов.
В определенный момент система клиренса не поспевает за продукцией, достигается критический уровень концентрации метаболитов, что приводит к торможению нейронов, ответственных за бодрствование, до достижения определенных концентраций накопленных метаболитов. Человек чувствует себя вялым.
К главным метаболитам, с которыми связывают инициацию сна, относят аденозин. Образуясь в ходе обычного энергетического обмена клеток, аденозин взаимодействует с метаботропными рецепторами (A1 и A2) на мембранах нейронов, вызывая торможение последних.
Было показано наличие А1-рецепторов в области нейронов базального ядра переднего мозга, которая наравне с глутаматергической системой покрышки моста являет собой важнейшую часть восходящей активирующей системы (ascending arousal system, AAS).
Однако эксперименты на крысах поставили под сомнение роль А1-рецепторов на нейронах BF в регуляции медленного сна.
Помимо этого, группой японских ученых была показана роль простагландина D2 в активации аденозинергических нейронов — https://vk.cc/6T1ae3, что может играть немалую роль в развитии гиперсомнии у некоторых лиц с ЧМТ или воспалением мозговых оболочек. Поэтому роль аденозина до сих пор остается не раскрытой.
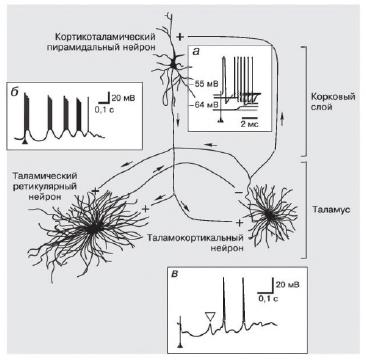
После инициации сна подключается активный механизм его поддержания. На ЭЭГ мы увидим сонные веретёна и К-комплексы. Основную роль в формировании сонных веретён играет таламус, который содержит 2 функциональных типа нейронов: релейные и ретикулярные.
В процессе бодрствования релейные таламические нейроны находятся в “stand by” режиме и готовы к передаче большинства сенсорных и внутренних сигналов, поступающих в кору больших полушарий. Активность этих нейронов поддерживается “системой бодрствования”, но все же главную роль в активации таламуса отдают глутаматергической системе покрышки моста (LDT/PPT).
В процессе инициации сна, когда происходит активация VLPO-нейронов, происходит одновременное снижение возбудимости релейных нейронов и активация ретикулярных. Ретикулярные таламические нейроны через стимуляцию постсинаптических ГАМК-В рецепторов ингибируют релейные нейроны путем их гиперполяризации.
Что происходит с потоком информации в этот момент? Поступающие с периферии сенсорные сигналы блокируются в таламусе и кора полушарий «лишается» информации об окружающем мире, переключаясь тем самым на внутренние сигналы за счет наличия на мембране корковых нейронов в области дендритов специфических низкопороговых Ca2+ — каналов, которые открываются в процессе гиперполяризации мембраны, вызывая деполяризацию самих нейронов.
Сонные веретёна представляют собой постингибиторные рикошетные взрывы потенциалов действия (postinhibitory rebound spike bursts, PRSB), возникающие на ретикулярных таламических, таламокортикальных и, собственно, кортикоталамических пирамидных нейронах, которые имеют обширные двусторонние связи — трехнейронная дуга Стериаде.
Дельта волны, характеризующие 3 и 4 фазы медленного сна, формируются в неокортексе за счет активации гиперполяризацией HCN — каналов (funny — токи) и и низкопороговых Са2+ — каналов кортико-кортикальных и кортикоталамических пирамидных клеток.
В среднем активность нейронов на протяжении медленного сна снижена, протекают процессы спраутинга (интенсивного ветвления аксонов), удаления функционально старых синаптических контактов, однако усиленно протекает синтез новых макромолекул и происходит восстановление гомеостаза клеток мозга.
Подводя итог, можно сказать, что роль NREM — сна чисто восстановительная. Вместе с процессом бодрствования, медленный сон поддерживает мозговой гомеостаз, а также обеспечивает нормальное функционирование таламо-кортикальной системы.
Телеграм: t.me/ainewsline
Источник: vk.cc