Клики нейронов, связанных с полостями, обеспечивают отсутствие связи между структурой и функцией
МЕНЮ
Главная страница
Поиск
Регистрация на сайте
Помощь проекту
Архив новостей
ТЕМЫ
Новости ИИ
Городские сумасшедшие
ИИ в медицине
ИИ проекты
Искусственные нейросети
Искусственный интеллект
Слежка за людьми
Угроза ИИ
ИИ теория
Компьютерные науки
Машинное обуч. (Ошибки)
Машинное обучение
Машинный перевод
Нейронные сети начинающим
Психология ИИ
Реализация ИИ
Реализация нейросетей
Создание беспилотных авто
Трезво про ИИ
Философия ИИ
Генетические алгоритмы
Капсульные нейросети
Основы нейронных сетей
Промпты. Генеративные запросы
Распознавание лиц
Распознавание образов
Распознавание речи
Творчество ИИ
Техническое зрение
Чат-боты
Авторизация
2017-06-18 14:40





Как и следовало ожидать, большая часть этого препринта про клики в направленных графах, а не про нейроны как таковые. Это к тому, что именно материал препринта состоит из алгебраических доказательств, оторванных от предметной области как таковой.]
Отсутствие формальной связи между структурой нейронной сети и ее возникающей функцией затрудняет наше понимание того, как мозг обрабатывает информацию. Мы подошли ближе к описанию такой связи, учитывая направление синаптической передачи, построим графики сети, которые отражают направление потока информации, и анализируя эти направленные графики с использованием алгебраической топологии. Применение этого подхода к локальной сети нейронов в неокортексе показало удивительно сложную и ранее невидимую топологию синаптической связи. Синаптическая сеть содержит обилие кликов нейронов, связанных в полостях, которые приводят к возникновению коррелированной активности. В ответ на раздражители коррелированная активность связывает синаптически связанные нейроны с функциональными кликами и полостями, которые эволюционируют в стереотипной последовательности до максимальной сложности. Мы предлагаем, чтобы мозг стимулировал стимулы, создавая все более сложные функциональные клики и полости.
1. Введение
Как структура сети определяет ее функцию, не совсем понятна. В частности, для нейронных сетей нам не хватает единой математической основы для однозначного описания возникающего поведения сети с точки зрения ее базовой структуры (Bassett and Sporns, 2017). Хотя теория графов использовалась для анализа топологии сети с некоторым успехом (Bullmore and Sporns, 2009), текущие методы обычно ограничиваются анализом того, как локальная связь влияет на локальную активность (Pajevic and Plenz, 2012; Chambers and MacLean, 2016) или глобальной сети (Hu et al., 2014) или как глобальные сетевые свойства, такие как связь и баланс возбуждающих и тормозных нейронов, влияют на динамику сети (Renart et al., 2010; Rosenbaum et al., 2017). Одно из таких свойств глобальной сети - это мир. Хотя было показано, что малый мир оптимизирует обмен информацией (Latora and Marchiori, 2001) и что адаптивная переадресация во время хаотической деятельности приводит к созданию сетей малого мира (Gong and Leeuwen, 2004), степень малости не может описывать большинство местных Свойства сети, такие как разные роли отдельных нейронов.
Алгебраическая топология (Munkres, 1984) предлагает уникальное преимущество предоставления методов для количественного описания свойств локальной сети и глобальных свойств сети, которые возникают из локальной структуры, тем самым объединяя оба уровня. Совсем недавно алгебраическая топология была применена к функциональным сетям между областями мозга с использованием fMRI (Petri et al., 2014) и между нейронами, использующими нейронную активность (Giusti et al., 2015), но основные синаптические связи (структурная сеть) были неизвестны , Более того, все формальные топологические анализы упускают из виду направление потока информации, поскольку они анализируют только неориентированные графики.
Мы разработали математическую основу для анализа структурной и функциональной топологии сети, интеграции локальных и глобальных описаний, что позволяет нам установить четкую взаимосвязь между ними. Мы представляем сеть в виде ориентированного графа с нейронами как вершинами и синаптическими связями, направленными от до-постсинаптических нейронов в виде ребер, которые могут быть проанализированы с использованием элементарных инструментов из алгебраической топологии (Munkres, 1984). Структурный график содержит все синаптические соединения, в то время как функциональный граф является подграфом структурного графа, содержащего только те соединения, которые активны в определенном временном ящике (т. е. Когда постсинаптический нейрон срабатывает в течение короткого периода пресинаптического всплеска ). Затем ответ на стимул может быть представлен и изучен как временной ряд функциональных графов.
Сети часто анализируются в терминах групп узлов, которые связаны друг с другом, называются кликами. Число нейронов в клике определяет его размер или, более формально, его размерность. В ориентированных графах естественно рассматривать направленные клики, которые представляют собой клики, содержащие один источник нейронов и единственный нейронный нейрон и отражающий конкретный мотив связности (Song et al., 2005; Perin et al., 2011), где поток Информации через группу нейронов имеет недвусмысленное направление. Способ, которым целенаправленные связки связывают друг с другом, может быть представлен геометрически. Когда направленные клики связываются надлежащим образом, разделяя нейроны и не образуя большей клики из-за недостающих соединений, они образуют в этом геометрическом представлении полости («дырки», «пустоты») с высокоразмерными полостями, образующимися при больших (больших) Клики соединяются вместе. Направленные клики описывают поток информации в сети на местном уровне, а полости обеспечивают глобальную меру потока информации во всей сети. Используя эти естественно возникающие структуры, мы установили прямую связь между структурным графом и появляющимся потоком информации в ответ на раздражители, захваченные через временные ряды функциональных графов.
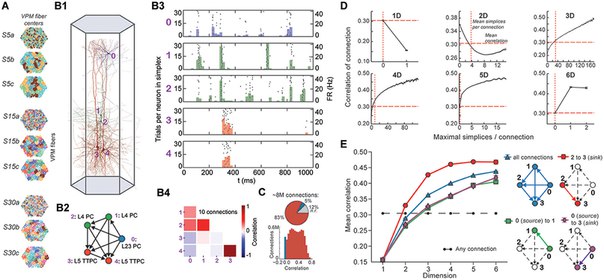
Мы применили эту схему для цифровых реконструкций неокортечной микросхемы крысы, которые очень напоминают биологическую ткань с точки зрения количества, типов и плотности нейронов и их синаптической связности (модель «микросоединителя» для кортикальной микросхемы, рисунки 1А, В, см. Markram et al., 2015; Reimann et al., 2015). Моделирование восстановленной микросхемы воспроизводило множественные возникающие электрические поведения, обнаруженные экспериментально в неокортексе (Markram et al., 2015). Микросхема, образованная ~ 8 млн соединений (ребер) между ~ 31 000 нейронов (вершин), была восстановлена из экспериментальных данных, руководствуясь биологическими принципами организации и итерационно уточнена до тех пор, пока не будет подтверждена батарея независимых анатомических и физиологических данных, полученных из экспериментов , Несколько экземпляров реконструкции обеспечивают статистический и биологический диапазон микросхем для анализа.
Мы обнаружили удивительно большое количество и разнообразие высокоразмерных направленных клик и полостей, которые ранее не встречались в нейронных сетях, как биологических, так и искусственных, и в гораздо большем количестве, чем в различных нулевых моделях направленных сетей. Топологические метрики, отражающие количество направленных клик и полостей, не только отличали реконструкцию от всех нулевых моделей, но также выявляли тонкие различия между реконструкциями на основе биологических наборов данных от разных животных, что указывало на то, что индивидуальные вариации биологической детали неокортикальных микросхем отражаются в репертуаре Направленных клик и полостей. Когда мы моделировали активность микросхем в ответ на сенсорные раздражители, мы наблюдали, что парные корреляции в активности нейронов увеличивались с числом и размером направленных клик, которым принадлежит пара нейронов, что указывает на то, что иерархическая структура сети формирует иерархию коррелированных Мероприятия. Фактически, мы обнаружили иерархию коррелированной активности между нейронами даже внутри одной направленной клики. Во время активности формировалось гораздо больше высокоразмерных направленных клик, чем следовало ожидать от числа активных соединений, что дополнительно указывало на то, что коррелированная активность имеет тенденцию связывать нейроны в высокоразмерные активные клики.
После применения пространственно-временного стимула к сети мы обнаружили, что при коррелированной активности активные клики образуют все более высокоразмерные полости (т.е. Полости, образованные все более крупными кликами). Более того, мы обнаружили, что в то время как разные пространственно-временные стимулы, применяемые к одной и той же схеме, и один и тот же стимул, применяемый к различным схемам, производят разные формы активности, все они проявляют ту же общую эволюцию, где формальные связи между все более высокоразмерными кликами формируются, а затем распадаются ,
2. Результаты
2.1. Случай для ориентированных упрощений
Сети нейронов, соединенных электрическими синапсами (щелевые соединения), могут быть представлены как неориентированные графики, где информация может течь в обоих направлениях. Сети с химическими синапсами, которые налагают одно направление синаптической связи от пресинаптического нейрона (рис. 1В2, В3), более точно представлены в виде ориентированных графов. Экспериментальные сети субсэмплирования нейронов выявили небольшие мотивы синаптической связи, но не большие клики нейронов (Song et al., 2005; Perin et al., 2011). Зная полную направленную сеть нейронов, как и в случае восстановленной микросхемы, позволяет обнаружить все клики, направленные и другие (рис. 1).
Когда направление соединений не принимается во внимание, теряется большая информация. Например, в неориентированном случае существует только одна возможная конфигурация для клики четырех полностью связанных нейронов (рис. 2A1, слева). Однако в рассматриваемом случае существует 36 = 729 возможных конфигураций, так как каждое из шести соединений может находиться в одном из трех состояний (i ? j, j ? i или i ? j типов соединений, рис. 2A1 справа).
Клика с обратными соединениями содержит два или более клика, состоящих только из однонаправленных соединений (рис. 2A2). Когда рассматриваются только однонаправленные соединения, существует 26 возможных конфигураций четырех полностью связанных нейронов, которые имеют два типа: те, которые содержат циклы (40 конфигураций, рис. 2А3 слева; раздел 4.1.3), а также те, которые этого не делают (24 Конфигурации; Рисунок 2A3 справа). Направленные клики - это именно ациклические клики. Чистая направленность информационного потока через любой мотив может быть определена как сумма по всем нейронам квадратов различий между их степенью и их степенью (см. Уравнение 2, рис. S1). Направленные клики имеют наивысшую чистую направленность среди всех клик (рис. S1, раздел 4.1.4). Клика, содержащая циклы, всегда разлагается в направленные клики с тем же числом нейронов или меньше, по крайней мере любая единственная связь между двумя нейронами образует 2-клику. Таким образом, циклическая клика трех нейронов распадается на три 2-клика. Следуя соглашениям в алгебраической топологии, мы ссылаемся на направленные клики n нейронов в качестве ориентированных симплексов размерности n-1 или направленных (n-1) -симплексов (которые отражают их естественное геометрическое представление как (n-1) -мерных многогранников) ( См. Рисунок S2, раздел 4.1.3). Соответственно, их подклассы называются субакрементами.
2.2. Изобилие управляемых упрощений
2.2.1. Реконструированная неокортечная микросхема
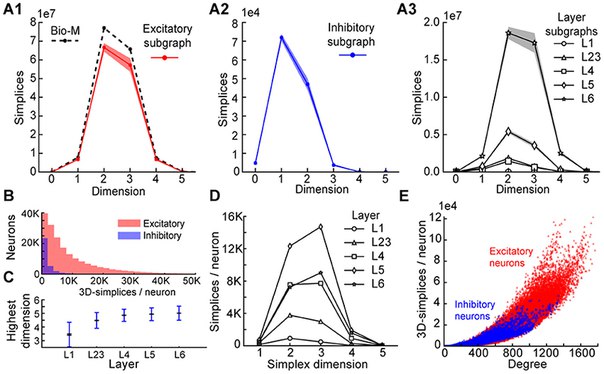
Мы проанализировали 42 варианта восстановленного микросоединения, сгруппированных в шесть наборов, каждый из которых состоял из семи статистически изменяющихся экземпляров (Markram et al., 2015; Section 4.3). Первые пять наборов были основаны на конкретных высотах шести слоев неокортекса, плотности клеток и распределениях различных типов клеток, экспериментально измеренных у пяти разных крыс (Bio1-5), в то время как шестое представляет среднее из этих измерений (Bio- М). Индивидуальные экземпляры внутри набора варьировались в зависимости от результата стохастических частей процесса реконструкции. Удивительно, но мы обнаружили, что в реконструкциях последовательно содержались ориентированные симплексы размеров до 6 или 7, с 80 миллионами направленных 3-симплексов (рис. 2B, синий). Это первый признак существования такого огромного числа высокоразмерных направленных симплексов в неоконтикальной микросхеме или в любой нейронной сети.
2.2.2. Модели управления
Чтобы сравнить эти результаты с нулевыми моделями, мы рассмотрели, как числа направленных симплексов в этих реконструкциях отличались от числа искусственных схем и схем, в которых некоторые биологические правила связности были опущены (см. Раздел 4.4). Для одного элемента управления мы сгенерировали пять случайных графов Erd?s-R?nyi (ER) равного размера (~ 31 000 вершин) и ту же среднюю вероятность соединения, что и схема Bio-M (~ 0,8%; ~ 8 миллионов ребер) (рисунок 2B; темный зеленый). Для другого мы построили схему с теми же трехмерными нейронами модели, что и схема Bio-M, но связали нейроны с использованием правила случайной связи [«Peters's Rule» (Peters and Feldman, 1976), PR; Рисунок 2B, красный]. Для последнего контроля мы связывали нейроны в цепи Bio-M в соответствии с вероятностными вероятностями соединения между различными морфологическими типами нейронов. Поскольку этот контроль аналогичен выводу соединения из среднего перекрытия нейронных беседок (Shepherd et al., 2005), он сохраняет общие биологические (GB) особенности связи между различными типами нейронов (Reimann et al., 2015), исключая Только явная попарная связь между отдельными нейронами, которая определяется перекрытием их конкретных беседок (рис. 2B, желтый). Во всех случаях количество направленных симплексов размером более 1 было намного меньше, чем в цепи Bio-M. Кроме того, относительные различия между Bio-M и нулевыми моделями заметно увеличились с измерением.
Телеграм: t.me/ainewsline
Источник: vk.com