Нейробиология и искусственный интеллект: часть первая — ликбез
МЕНЮ
Главная страница
Поиск
Регистрация на сайте
Помощь проекту
Архив новостей
ТЕМЫ
Новости ИИ
Голосовой помощник
Городские сумасшедшие
ИИ в медицине
ИИ проекты
Искусственные нейросети
Искусственный интеллект
Слежка за людьми
Угроза ИИ
Разработка ИИГородские сумасшедшие
ИИ в медицине
ИИ проекты
Искусственные нейросети
Искусственный интеллект
Слежка за людьми
Угроза ИИ
Атаки на ИИ
ИИ теория
Компьютерные науки
Машинное обуч. (Ошибки)
Машинное обучение
Машинный перевод
Нейронные сети начинающим
Психология ИИ
Реализация ИИ
Реализация нейросетей
Создание беспилотных авто
Трезво про ИИ
Философия ИИ
Внедрение ИИИИ теория
Компьютерные науки
Машинное обуч. (Ошибки)
Машинное обучение
Машинный перевод
Нейронные сети начинающим
Психология ИИ
Реализация ИИ
Реализация нейросетей
Создание беспилотных авто
Трезво про ИИ
Философия ИИ
Big data
Генетические алгоритмы
Капсульные нейросети
Основы нейронных сетей
Промпты. Генеративные запросы
Распознавание лиц
Распознавание образов
Распознавание речи
Творчество ИИ
Техническое зрение
Чат-боты
Работа разума и сознаниеМодель мозгаРобототехника, БПЛАТрансгуманизмОбработка текстаТеория эволюцииДополненная реальностьЖелезоКиберугрозыНаучный мирИТ индустрияРазработка ПОТеория информацииМатематикаЦифровая экономика
Генетические алгоритмы
Капсульные нейросети
Основы нейронных сетей
Промпты. Генеративные запросы
Распознавание лиц
Распознавание образов
Распознавание речи
Творчество ИИ
Техническое зрение
Чат-боты
Авторизация
2017-01-21 10:50

Так уж получилось, что я уже давненько читаю Хабр и особенно раздел про искусственный интеллект, однако до сих пор так и не отважился внести свою лепту в общий уровень энтропии.
Оживленные дискуссии в комментариях показывают живой интерес к теме и одновременно большое разнообразие точек зрения, мнений и уровней подготовки. Просмотрев историю публикаций, я как-то не нашел важного отправного пункта для многих рассуждений, а именно – какого-нибудь описания механизмов передачи сигналов в мозге. Те, кто пишут про нейронные сети и прогресс в компьютерных моделях интеллекта обычно вскользь упоминают про синапсы и медиаторы (что вполне достаточно для их целей), те же, кто пытается понять природу естесственного интеллекта в основном рассуждают философскими категориями. В результате, множество коментариев содержат отсылки к популярным домыслам и мифам, не находящим подтверждения в современных исследованиях.
В данной статье я в сжатой форме попытаюсь дать ответы на следующие вопросы:
— что же такое нейрон, как он устроен и работает?
— что происходит в синапсах, когда нейроны общаются друг с другом?
А в следующей (-щих):
— как интеллект и сознание связаны с активностью нейронов? (здесь же про то, как информация обрабатывается мозгом, нейропластичность, квантовую теорию сознания, сон и т.д.)
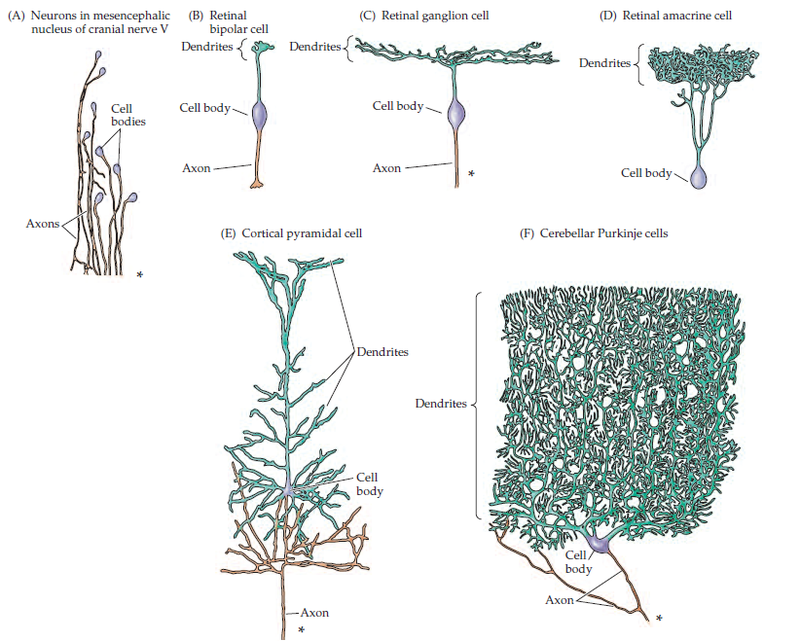
Рис. 1. Различные формы нейронов. Здесь и далее картинки взяты из “Neuroscience”, 3d edition, Dale Purves et al.
Во-первых, стоит упомянуть, что в человеческом организме далеко не один вид нейронов, однако при всех их различиях (рис. 1) у них есть много общего, в том числе и с точки зрения механизмов обеспечения их прямой функциональности. Во-вторых, в мозгу кроме нейронов есть еще множетсво вспомогательных клеток – клеток нейроглии, или просто глии. Этих клеток в среднем в 3 раза больше чем нейронов, и они обеспечивают нейроны питанием, энергией и необходимыми веществами. В последнее время им стало уделяться все больше внимания в силу того, что было показано их влияние на работу нейронов (например [1]) в области обучения и памяти. Здесь стоило бы отмтетить, что являясь клетками поддержки, нейроглия не может не влиять на работу нейронов, ведь голодный нейрон не может работать так же, как и сытый, однако не стоит переоценивать их роль в обработке информации: да, они влияют на работу нейронов и синапсов, но не определяют ее. Отсюда и пословица: сытый голодного не разумеет.
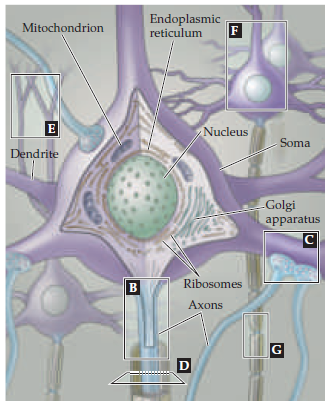 Рис. 2. Схематичное изображение структуры нейрона.
Рис. 2. Схематичное изображение структуры нейрона.
Но вернемся к нейронам и сигналам, которые они продуцируют. Абстрактная усредненная структура нейрона предс тавляет собой что-то типа изображенного на рис. 2. и состоит из тела клетки (soma), в котором можно найти все элементы обычной живой клетки (ядро, аппарат Гольджи, митохондрии, т.д.), дендритов – отростков, служащих как инпут от других нейронов, и обычно одного длинного аксона – по которому нейрон вещает свое мнение другим нейронам. Из этой структуры и пошли быть все математические модели перцептронов – много входов, черный ящик, один выход, профит. А что же происходит в черном ящике? Как нейрон обрабатывает входящие сигналы и на чем основаны математические модели нейронов? Оказывается, не все так сложно, как могло бы быть. Рассмотрим, например, механизм работы аксона по передаче сигнала.
 Рис. 3. Структура аксона с основными действующими элементами для распространения сигнала: протеинами, отвечающими за перенос ионов из внутренней части клетки наружу и наоборот.
Рис. 3. Структура аксона с основными действующими элементами для распространения сигнала: протеинами, отвечающими за перенос ионов из внутренней части клетки наружу и наоборот.
Как видно из рисунка 3, аксон представляет собой обычную клеточную мембрану, состоящую из двойного слоя липидов, перемежающихся различными протеинами, плюс, то что не показано на картинке – цитоскелет, состоящий из протеиновых микро-трубочек и соединительных протеинов. Тут важно заметить, что внутри и снаружи аксона химический состав жидкостей различен. И, как мы увидим позже, очень важную роль играют градиенты концентраций ионов натрия, калия, кальция и хлора. Различная концентрация ионов по обе стороны мембраны активно поддерживается как раз теми протеинами, которые в мембране и находятся в большом количестве. Часть из них поддерживает определенный потенциал мембраны, так называемый resting potential (RP): у различных организмов он варьируется от -40 до -90 mV. Получается он благодаря тому, что специальные протеины (их называют active transporters) качают в одностороннем порядке ионы против градиента концентрации, в то время как другие протеины (называемые ion channels) позволяют определенным ионам течь в обратном направлении. Равновесие этих двух процессов влияет как на RP, так и на action potential (AP) – основной сигнал передаваемый от одного нейрона к другому, а так же на синаптический и рецепторный потенциалы, которые и обеспечивают АР.
Итак, поскольку АР это то, что передается от одного нейрона к другому, для нас он будет наиболее важен, посему давайте рассмотрим его более детально. Работа протеинов по перекачке ионов и контролю уровня RP в определенных пределах довольно хорошо организована и даже помехоустойчива. Это можно продемонстрировать, прикладывая определенные потенциалы к мембране (рис. 4).
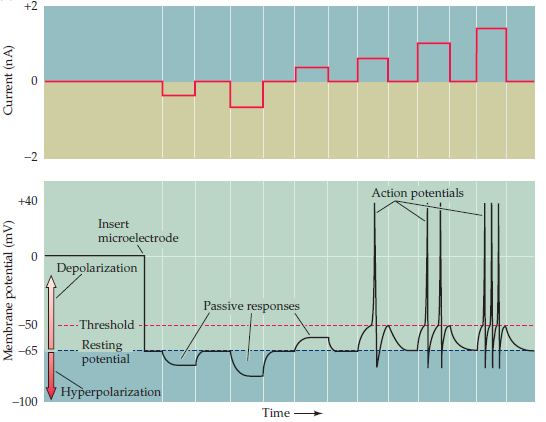 Рис. 4. Схематичное изображение условий генерации пассивных и активных сигналов.
Рис. 4. Схематичное изображение условий генерации пассивных и активных сигналов.
Как видно из диаграммы, при равновесном значении RP ток через мембрану не течет. Если к мембране приложить внешний потенциал, то в определенных пределах ее отклик будет линейным. Однако, по пересечению некоего порогового значения (Threshold) происходит «взрывная» деполяризация мембраны – тот самый action potential. Причем важно заметить: прикладывая больший потенциал к мембране, АР не становится больше или длиннее, но частота следования становится больше! И тут никакого волшебства: в спокойное время, натриевые каналы закрыты и градиент концентрации натрия обеспечивается ион-транспортными протеинами. Однако при пересечении порогового значения потенциала мембраны натриевые каналы включаются и очень быстро пропускают большое количество ионов внутрь клетки, быстро изменяя общий потенциал мембраны. Калиевые каналы такого обращения терпеть не будут и тоже откроются, но равновесный градиент концентраций калия противоположен градиенту концентраций натрия, поэтому калий потечет из аксона наружу, тем самым нейтрализуя эффект вызванным натрием и восстанавливая равновесное значение RP. Благодаря разной скорости протекания этих двух процессов на короткое время образуется чрезмерно отрицательный потенциал мембраны, но он компенсируется работой ион-транспортых протеинов.
Так что фактически аксон – простой АЦП, который кодирует амплитуду входящего сигнала частотой выходящих сигналов, плюс некий порог ниже которого он вообще не напрягается, и передает этот сигнал дальше. Распространение сигнала вдоль аксона регулируется все теми же протеинами: когда в одной части аксона появляется АР, он меняет концентрации ионов в некоей окрестности, и там происходит то же самое, только с определенной временной задержкой, отсюда вполне конечная скорость распространения нервных импульсов: от 0.5-10 м/с по коротким нейронам до 150 м/с по длинным аксонам, окруженным миелином.
Начать, наверное, стоит с того, что упомянуть что синапс это соединение аксона одного нейрона с дендритом другого и синапсы бывают двух видов: электрические и химические. Первые довольно редки, но фактически представляют возможность передавать электрический сигнал напрямую от одного нейрона к другому. Это происходит тогда, когда синаптическое соединение двух нейронов настолько близкое, что они «сливаются» и ионные каналы одного нейрона напрямую соединены с ионными каналами другого нейрона, позволяя маленьким молекулам перемещаться между ними. Таким образом электрические синапсы просто позволяют ионным токам перетекать между нейронами. Интересная особенность такого соединения – оно работает в обоих направлениях. Другая особенность – они чрезвычайно быстрые. Поэтому такие синапсы используются в популяциях нейронов, которые должны быть хорошо синхронизированы, например которые генерируют ритмическую активность для дыхания.
 Рис. 5. Полный цикл передачи сигнала через химический аксон: 1 – синтез и хранение трансмиттера; 2 – приход АР; 3 – открытие кальциевых каналов под воздействием АР; 4 – перетекание ионов кальция через открытый канал; 5 – слияние пузырьков с трансмиттером с мембраной под воздействием излишней концентрации ионов кальция; 6 – высвобождение трансмиттера в синаптическое пространство; 7 – связывание трансмиттера с рецепторами на дендритной стороне синапса; 8 – открытие ионных каналов под воздействием трансмиттера; 9 – образование АР под воздействием тока ионов через каналы; 10 – возвращение пузырьков для наполнения новым трансмиттером.
Рис. 5. Полный цикл передачи сигнала через химический аксон: 1 – синтез и хранение трансмиттера; 2 – приход АР; 3 – открытие кальциевых каналов под воздействием АР; 4 – перетекание ионов кальция через открытый канал; 5 – слияние пузырьков с трансмиттером с мембраной под воздействием излишней концентрации ионов кальция; 6 – высвобождение трансмиттера в синаптическое пространство; 7 – связывание трансмиттера с рецепторами на дендритной стороне синапса; 8 – открытие ионных каналов под воздействием трансмиттера; 9 – образование АР под воздействием тока ионов через каналы; 10 – возвращение пузырьков для наполнения новым трансмиттером.
Химические синапсы работают немного по-другому (рис. 5). Вкратце, когда приходит АР, со стороны аксона выпускается нейротрансмиттер, который диффундирует сквозь синаптическое пространство и связывается с рецепторами на стороне дендрита, активируя их. Активированные рецепторы деполяризуют мембрану дендрита и тем самым включают процесс распростанения АР по своей мембране. В теории просто, а на практике вяснилось, что нейротрасмиттеров под 100 различных видов. Спрашивается, а зачем столько? Исследования показали, что многие нейроны вырабатывают одновременно несколько неротрансмиттеров, которые, в свою очередь, по-разному влияют на рецепторы пост-синаптической (принимающей сигнал) мембраны. Например, нейротрансмиттеры небольшого размера, выделяющиеся обычно при редко приходящих АР, быстро перетекают через синаптическое пространство и вызывают быструю реакцию, в то время как большие молекулы, выделяющиеся при часто приходящих АР, перемещаются дольше, но и связываются с рецепторами сильнее, тем самым порождая более длительную деполяризацию мембраны. Все это создает предпосылки для более качественной передачи пришедшего сигнала. Так же, важно отметить, что одни нейротрансмиттеры могут возбуждать принимающий нейрон, а другие – наоборот ингибировать возбуждение.
Вот как-то так.
Пока что про искусственный интеллект не особо много получилось, но и до него дойдем.
— [1] гуглим «mystery-of-the-human-brains-glia-cells-solved-key-to-learning-information-processing»
Оживленные дискуссии в комментариях показывают живой интерес к теме и одновременно большое разнообразие точек зрения, мнений и уровней подготовки. Просмотрев историю публикаций, я как-то не нашел важного отправного пункта для многих рассуждений, а именно – какого-нибудь описания механизмов передачи сигналов в мозге. Те, кто пишут про нейронные сети и прогресс в компьютерных моделях интеллекта обычно вскользь упоминают про синапсы и медиаторы (что вполне достаточно для их целей), те же, кто пытается понять природу естесственного интеллекта в основном рассуждают философскими категориями. В результате, множество коментариев содержат отсылки к популярным домыслам и мифам, не находящим подтверждения в современных исследованиях.
В данной статье я в сжатой форме попытаюсь дать ответы на следующие вопросы:
— что же такое нейрон, как он устроен и работает?
— что происходит в синапсах, когда нейроны общаются друг с другом?
А в следующей (-щих):
— как интеллект и сознание связаны с активностью нейронов? (здесь же про то, как информация обрабатывается мозгом, нейропластичность, квантовую теорию сознания, сон и т.д.)
Так что же такое нейрон, как он устроен и работает?
Рис. 1. Различные формы нейронов. Здесь и далее картинки взяты из “Neuroscience”, 3d edition, Dale Purves et al.
Во-первых, стоит упомянуть, что в человеческом организме далеко не один вид нейронов, однако при всех их различиях (рис. 1) у них есть много общего, в том числе и с точки зрения механизмов обеспечения их прямой функциональности. Во-вторых, в мозгу кроме нейронов есть еще множетсво вспомогательных клеток – клеток нейроглии, или просто глии. Этих клеток в среднем в 3 раза больше чем нейронов, и они обеспечивают нейроны питанием, энергией и необходимыми веществами. В последнее время им стало уделяться все больше внимания в силу того, что было показано их влияние на работу нейронов (например [1]) в области обучения и памяти. Здесь стоило бы отмтетить, что являясь клетками поддержки, нейроглия не может не влиять на работу нейронов, ведь голодный нейрон не может работать так же, как и сытый, однако не стоит переоценивать их роль в обработке информации: да, они влияют на работу нейронов и синапсов, но не определяют ее. Отсюда и пословица: сытый голодного не разумеет.
Рис. 2. Схематичное изображение структуры нейрона.Но вернемся к нейронам и сигналам, которые они продуцируют. Абстрактная усредненная структура нейрона предс тавляет собой что-то типа изображенного на рис. 2. и состоит из тела клетки (soma), в котором можно найти все элементы обычной живой клетки (ядро, аппарат Гольджи, митохондрии, т.д.), дендритов – отростков, служащих как инпут от других нейронов, и обычно одного длинного аксона – по которому нейрон вещает свое мнение другим нейронам. Из этой структуры и пошли быть все математические модели перцептронов – много входов, черный ящик, один выход, профит. А что же происходит в черном ящике? Как нейрон обрабатывает входящие сигналы и на чем основаны математические модели нейронов? Оказывается, не все так сложно, как могло бы быть. Рассмотрим, например, механизм работы аксона по передаче сигнала.
Рис. 3. Структура аксона с основными действующими элементами для распространения сигнала: протеинами, отвечающими за перенос ионов из внутренней части клетки наружу и наоборот.1. Структура аксона
Как видно из рисунка 3, аксон представляет собой обычную клеточную мембрану, состоящую из двойного слоя липидов, перемежающихся различными протеинами, плюс, то что не показано на картинке – цитоскелет, состоящий из протеиновых микро-трубочек и соединительных протеинов. Тут важно заметить, что внутри и снаружи аксона химический состав жидкостей различен. И, как мы увидим позже, очень важную роль играют градиенты концентраций ионов натрия, калия, кальция и хлора. Различная концентрация ионов по обе стороны мембраны активно поддерживается как раз теми протеинами, которые в мембране и находятся в большом количестве. Часть из них поддерживает определенный потенциал мембраны, так называемый resting potential (RP): у различных организмов он варьируется от -40 до -90 mV. Получается он благодаря тому, что специальные протеины (их называют active transporters) качают в одностороннем порядке ионы против градиента концентрации, в то время как другие протеины (называемые ion channels) позволяют определенным ионам течь в обратном направлении. Равновесие этих двух процессов влияет как на RP, так и на action potential (AP) – основной сигнал передаваемый от одного нейрона к другому, а так же на синаптический и рецепторный потенциалы, которые и обеспечивают АР.
2. Что такое сигнал для аксона и откуда он берется?
Итак, поскольку АР это то, что передается от одного нейрона к другому, для нас он будет наиболее важен, посему давайте рассмотрим его более детально. Работа протеинов по перекачке ионов и контролю уровня RP в определенных пределах довольно хорошо организована и даже помехоустойчива. Это можно продемонстрировать, прикладывая определенные потенциалы к мембране (рис. 4).
Рис. 4. Схематичное изображение условий генерации пассивных и активных сигналов.Как видно из диаграммы, при равновесном значении RP ток через мембрану не течет. Если к мембране приложить внешний потенциал, то в определенных пределах ее отклик будет линейным. Однако, по пересечению некоего порогового значения (Threshold) происходит «взрывная» деполяризация мембраны – тот самый action potential. Причем важно заметить: прикладывая больший потенциал к мембране, АР не становится больше или длиннее, но частота следования становится больше! И тут никакого волшебства: в спокойное время, натриевые каналы закрыты и градиент концентрации натрия обеспечивается ион-транспортными протеинами. Однако при пересечении порогового значения потенциала мембраны натриевые каналы включаются и очень быстро пропускают большое количество ионов внутрь клетки, быстро изменяя общий потенциал мембраны. Калиевые каналы такого обращения терпеть не будут и тоже откроются, но равновесный градиент концентраций калия противоположен градиенту концентраций натрия, поэтому калий потечет из аксона наружу, тем самым нейтрализуя эффект вызванным натрием и восстанавливая равновесное значение RP. Благодаря разной скорости протекания этих двух процессов на короткое время образуется чрезмерно отрицательный потенциал мембраны, но он компенсируется работой ион-транспортых протеинов.
Так что фактически аксон – простой АЦП, который кодирует амплитуду входящего сигнала частотой выходящих сигналов, плюс некий порог ниже которого он вообще не напрягается, и передает этот сигнал дальше. Распространение сигнала вдоль аксона регулируется все теми же протеинами: когда в одной части аксона появляется АР, он меняет концентрации ионов в некоей окрестности, и там происходит то же самое, только с определенной временной задержкой, отсюда вполне конечная скорость распространения нервных импульсов: от 0.5-10 м/с по коротким нейронам до 150 м/с по длинным аксонам, окруженным миелином.
Что происходит в синапсах, когда нейроны общаются друг с другом?
Начать, наверное, стоит с того, что упомянуть что синапс это соединение аксона одного нейрона с дендритом другого и синапсы бывают двух видов: электрические и химические. Первые довольно редки, но фактически представляют возможность передавать электрический сигнал напрямую от одного нейрона к другому. Это происходит тогда, когда синаптическое соединение двух нейронов настолько близкое, что они «сливаются» и ионные каналы одного нейрона напрямую соединены с ионными каналами другого нейрона, позволяя маленьким молекулам перемещаться между ними. Таким образом электрические синапсы просто позволяют ионным токам перетекать между нейронами. Интересная особенность такого соединения – оно работает в обоих направлениях. Другая особенность – они чрезвычайно быстрые. Поэтому такие синапсы используются в популяциях нейронов, которые должны быть хорошо синхронизированы, например которые генерируют ритмическую активность для дыхания.
Рис. 5. Полный цикл передачи сигнала через химический аксон: 1 – синтез и хранение трансмиттера; 2 – приход АР; 3 – открытие кальциевых каналов под воздействием АР; 4 – перетекание ионов кальция через открытый канал; 5 – слияние пузырьков с трансмиттером с мембраной под воздействием излишней концентрации ионов кальция; 6 – высвобождение трансмиттера в синаптическое пространство; 7 – связывание трансмиттера с рецепторами на дендритной стороне синапса; 8 – открытие ионных каналов под воздействием трансмиттера; 9 – образование АР под воздействием тока ионов через каналы; 10 – возвращение пузырьков для наполнения новым трансмиттером.Химические синапсы работают немного по-другому (рис. 5). Вкратце, когда приходит АР, со стороны аксона выпускается нейротрансмиттер, который диффундирует сквозь синаптическое пространство и связывается с рецепторами на стороне дендрита, активируя их. Активированные рецепторы деполяризуют мембрану дендрита и тем самым включают процесс распростанения АР по своей мембране. В теории просто, а на практике вяснилось, что нейротрасмиттеров под 100 различных видов. Спрашивается, а зачем столько? Исследования показали, что многие нейроны вырабатывают одновременно несколько неротрансмиттеров, которые, в свою очередь, по-разному влияют на рецепторы пост-синаптической (принимающей сигнал) мембраны. Например, нейротрансмиттеры небольшого размера, выделяющиеся обычно при редко приходящих АР, быстро перетекают через синаптическое пространство и вызывают быструю реакцию, в то время как большие молекулы, выделяющиеся при часто приходящих АР, перемещаются дольше, но и связываются с рецепторами сильнее, тем самым порождая более длительную деполяризацию мембраны. Все это создает предпосылки для более качественной передачи пришедшего сигнала. Так же, важно отметить, что одни нейротрансмиттеры могут возбуждать принимающий нейрон, а другие – наоборот ингибировать возбуждение.
Вот как-то так.
Пока что про искусственный интеллект не особо много получилось, но и до него дойдем.
— [1] гуглим «mystery-of-the-human-brains-glia-cells-solved-key-to-learning-information-processing»
Телеграм: t.me/ainewsline
Источник: habrahabr.ru