Сперматогенез: через тернии к звездам
МЕНЮ
Искусственный интеллект
Поиск
Регистрация на сайте
Помощь проекту
ТЕМЫ
Новости ИИ
Голосовой помощник
Городские сумасшедшие
ИИ в медицине
ИИ проекты
Искусственные нейросети
Слежка за людьми
Угроза ИИ
Компьютерные науки
Машинное обуч. (Ошибки)
Машинное обучение
Машинный перевод
Реализация ИИ
Реализация нейросетей
Создание беспилотных авто
Трезво про ИИ
Философия ИИ
Генетические алгоритмы
Капсульные нейросети
Основы нейронных сетей
Распознавание лиц
Распознавание образов
Распознавание речи
Техническое зрение
Чат-боты
Авторизация
2018-12-04 16:05

На определенном этапе сперматогенеза созревающие половые клетки должны пройти сквозь один из самых прочных барьеров в организме — гемато-тестикулярный барьер. Горячие споры о том, как им это удается, кипели в научном мире в течение десятилетий. Двум исследователям из США удалось разгадать эту загадку.
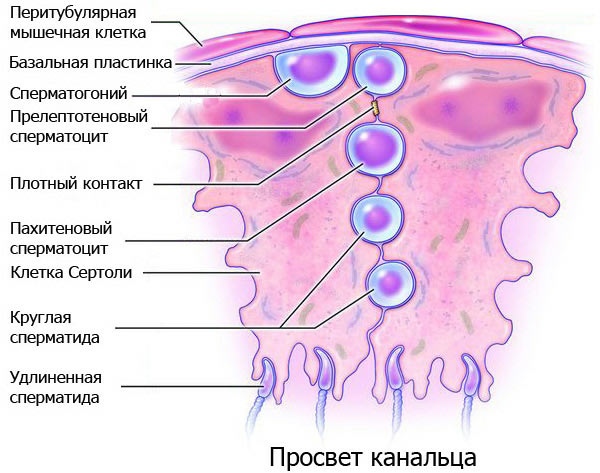
Сперматогенез — то есть, процесс созревания мужских половых клеток — это долгий путь постепенной дифференцировки, в результате которой из крупных, неподвижных стволовых клеток получаются маленькие, мобильные, терминально дифференцированные сперматозоиды. Схема сперматогенеза изображена на рисунке 1, и прежде, чем читать дальше, я советую читателю внимательно ее изучить.
Схема сперматогенеза
В этом процессе почти все клетки при делении не до конца расходятся, образуя синцитий, благодаря чему все физиологические процессы в них синхронизируются, и они могут одновременно совершать какие-то действия, например, двигаться или делиться. Сперматогенез можно разбить на такие стадии: 1) Все начинается с так называемых «недифференцированных сперматогониев». Они все обладают свойствами стволовых клеток [1]. Делятся они довольно вяло и безалаберно (то есть не в строго определенные моменты времени, а когда попало). Иногда они делают «шаг назад» по этой схеме, превращаясь в клетки предшествующих типов (например, два Apaired-сперматогония могут преспокойно поделиться и превратиться в пару Asingle) — это нужно, чтобы поддерживать популяцию стволовых клеток, а также регулировать итоговый выход сперматозоидов. 2) На следующей ступени находятся чуть более дифференцированные A1—4-сперматогонии. В отличие от своих предшественников (и так же, как и все последующие типы клеток), они делятся уже не когда попало, а «по часам», в строго определенные моменты времени, которые называются стадиями цикла семенного эпителия. Кроме того, на этом этапе наблюдается массовая гибель некоторых клонов клеток, необходимая для уменьшения итогового выхода спермы. И вот A4 дают при делении... 3) Еще более дифференцированные промежуточные и В-сперматогонии, при делении которых получаются... 4) Сильнодифференцированные сперматоциты первого порядка, которые вступают в мейоз и дают вначале сперматоциты второго порядка, а затем... 5) Сперматиды, которые не делятся, а только созревают, дифференцируются, из круглых становятся вытянутыми и наконец превращаются в... 6) Сперматозоиды, которые уплывают прочь, оставляя после себя так называемое остаточное тельце, состоящее из ненужной больше цитоплазмы. Рисунок автора по схемам из обзора [2] и классической книги [3].
Все описанные на рисунке 1 процессы происходят в семенных канальцах — тонких трубочках внутри мужских яичек. Представим себе такую трубочку в поперечном разрезе — получится кольцо. На самой наружной части этого кольца, базальной пластинке, вальяжно распластались As-сперматогонии. По мере дифференцировки, на каждом следующем этапе сперматогенеза, клетки (точнее, цепочки клеток, которые показаны на рис. 1) все сильнее сдвигаются оттуда ко внутренней части кольца (по направлению к его просвету), пока, наконец, почти дозревшие сперматиды не уплывают куда-то вдаль по этому просвету, по дороге окончательно созревая.
Созревающие половые клетки находятся в семенных канальцах не одни — помимо них там присутствуют так называемые клетки-няньки (nurse cells), которые поддерживают их, питают и защищают. Эти вспомогательные клетки называются клетками Сертоли, и когда мы говорим, что по мере дифференцировки половые клетки продвигаются от наружной части извитого канальца ко внутренней, это означает, что они движутся там сквозь «строй» клеток Сертоли (рис. 2).
И вот тут есть одна загвоздка. Дело в том, что проход через клетки Сертоли в одном месте накрепко заперт, и кажется, что пройти по нему невозможно.
Участок этот называется гемато-тестикулярным барьером (ГТБ). Он образован необычайно прочными межклеточными соединениями — плотными контактами (см. tight junction) — и располагается как раз там, где клетки окончательно перестают быть стволовыми и отправляются в необратимое путешествие по пути терминальной дифференцировки. То есть, сразу после того места, где сперматоциты первого порядка начинают готовиться к первому делению мейоза (поскольку они в этот момент находятся на стадии прелептотены, то называются прелептотеновыми сперматоцитами) и перед тем участком, где находятся пахитеновые (находящиеся на мейотической стадии пахитены) сперматоциты.
Зачем же необходим этот непреодолимый барьер? Как минимум по двум причинам.
Во-первых, для того, чтобы отделить стволовые клетки от дифференцированных. Дело в том, что для поддержания состояния «стволовости» клеткам необходимо особое микроокружение, так называемая «ниша» (см. stem cell niche) — «ареол» определенных веществ, которые не дают этим клеткам дифференцироваться и заставляют их быть стволовыми. Если в эту нишу попадут дифференцирующиеся клетки, то о всякой дифференцировке можно будет забыть: они начнут возвращаться в стволовое состояние и никогда уже не дадут начало сперматозоидам. Столь же печальны и перспективы стволовых клеток, попавших в микроокружение, необходимое для дифференцировки: они навсегда перестанут быть стволовыми, дифференцируются и в виде дозревающих на ходу сперматид уплывут вдаль по просвету канальца.
Во-вторых, и может быть, и в-главных, гемато-тестикулярный барьер необходим для того, чтобы ни в коем случае не допустить попадания дифференцирующихся половых клеток в кровь и в лимфу. Дело в том, что по мере дифференцировки на этих клетках появляется все больше и больше антигенов, и встреча их с антителами (которые в огромных количествах плавают в лимфе и крови) неминуемо приведет к аутоиммунной реакции, что может плохо кончиться как для бедных половых клеток, так и для организма в целом.
Итак, получается, что барьер этот крепок и непреодолим. И в то же время очевидно, что каким-то образом созревающим клеткам удается сквозь него пробираться. Как же это возможно?
Данный вопрос мучил умы ученых на протяжении многих десятилетий, пока, наконец, двое американских исследователей в серии простых, но элегантных экспериментов не расставили все по своим местам [4].
Использованная ими методика заключалась в следующем. Они брали семенные канальцы мышей, окрашивали их на белки, характерные только для плотных контактов, и белки, встречающиеся только в межклеточных мостиках клеточных клонов, а затем рассматривали получившиеся препараты под конфокальным микроскопом. Таким образом они могли рассмотреть взаимное расположение ГТБ и цепочек проходящих через него клеток.
Однако как же «поймать» клетки именно в тот момент, когда они преодолевают барьер? На первый взгляд, это можно сделать только путем долгих и скучных проб и ошибок, однако все гораздо проще, чем кажется. Дело в том, что сперматогенез в семенных канальцах проходит волнами, и каждая новая волна начинается тогда, когда предыдущая еще не закончилась. Иными словами, в тот момент, когда A1-сперматогонии начинают делиться где-то возле базальной пластинки, клетки предыдущей волны сперматогенеза находятся еще только на полпути к просвету канальца. Волны сперматогенеза синхронизированы между собой (это нужно, чтобы обеспечить стабильный и постоянный выход сперматозоидов), и поэтому, зная, где находятся клетки одной из волн сперматогенеза, можно точно сказать, где находятся клетки других волн. В данном случае исследователи выбирали те препараты, где почти дозревшие сперматиды покидали просвет канальца — это означало, что клетки одной из последующих волн сперматогенеза уже дошли до стадии прелептотеновых сперматоцитов и как раз собираются преодолевать гемато-тестикулярный барьер. (Узнать больше о стадиях цикла семенного эпителия и волнах сперматогенеза можно в классической статье [5].)
И выяснилась удивительная вещь. Оказалось, что барьер во время сперматогенеза не рвется. Вместо этого откуда-то возникает новый, второй слой барьера, в результате чего цепочки сперматоцитов (находящихся уже на стадии лептотены) окружаются барьером, как связки сосисок — полиэтиленовой упаковкой (рис. 3). Причем исследователям нигде не удалось увидеть, чтобы край какой-нибудь цепочки где-то прорывал один из слоев барьера — то есть, в любой момент прохождения сперматоцитов барьер остается целым и невредимым. Однако когда сперматоциты проходят область барьера, он снова становится однослойным.
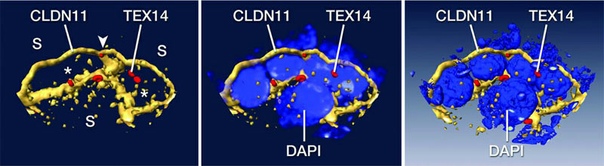
Подробнее разбираясь, каким же образом появляется новый слой барьера, ученые обнаружили, что большое значение в этом процессе играет белок клаудин 3, один из представителей обширного семейства клаудинов. Белки этого семейства являются одним из главных компонентов плотных контактов.
У каждого клаудина свой «характер» и своя роль при образовании или поддержании плотных контактов. И вот клаудин 3 нужен как раз при образовании новых контактов — в старых он практически не встречается. Так и в этот раз — исследователи показали, что новый слой барьера образуется при активном участии клаудина 3, однако по мере «старения» барьера клаудин 3 заменяется своим «родственником», клаудином 11, и, в конце концов, вовсе исчезает из плотных контактов (см. рис. 4).

Иными словами, картина выглядит примерно так. Цепочки сперматоцитов при своем движении к апикальной (обращенной в просвет) части канальца упираются в ГТБ. Тогда с базальной части этих сперматоцитов образуется новый, второй слой барьера, в результате чего сперматоциты оказываются внутри отдельного компартмента (той самой «сосисочной оболочки»). Двигаясь дальше, сперматоциты разрушают старый, апикальный слой, в результате чего новый, базальный слой остается единственным барьером — ну, до тех пор, конечно, пока к нему не подойдет новая партия сперматоцитов. То есть, получается, что при движении через него сперматоцитов барьер не разрушается и не сдвигается, а, можно сказать, самообновляется. Не правда ли, красивая картина возникает перед глазами?
Литература
- Ствол и ветки: стволовые клетки;
- de Rooij D.G., Griswold M.D. (2012). Questions about spermatogonia posed and answered since 2000. J. Androl. doi: 10.2164/jandrol.112.016832;См также: de Rooij D.G., Russell L.D. (2000). All you wanted to know about spermatogonia but were afraid to ask. J. Androl. 21, 776–799;;
- Bloom W., Fawcett D.W. (1975). Textbook of Histology. 10th Ed. Saunders, Philadelphia;
- Smith B.E., Braun R.E. (2012). Germ Cell Migration Across Sertoli Cell Tight Junctions. Science doi: 10.1126/science.1219969;
- Oakberg E.F. (1956). Duration of spermatogenesis in the mouse and timing of stages of the cycle of the seminiferous epithelium. Am. J. Anat. 99, 507–516;;
- Элементы: «Гемато-тестикулярный барьер не рвется, а обновляется»..
Источник: biomolecula.ru